The digestive system
Publisher Summary
This chapter focuses on the anatomy and histology of digestive system. The digestive system, or the gut, is a muscular tube lined with endoderm that traverses the body from mouth to anus, sealed at various points by muscular sphincters controlled by the autonomic nervous system. The digestive tube is subdivided into a number of organs on a basis of the anatomical and functional breaks as a result of the sphincters. The major part of the tube from esophagus to anus is made up of layers of muscle separated from each other by connective tissue in which lie blood vessels and nerves. The characteristic movement of the digestive tract is peristalsis. A peristaltic wave is a wave of contraction originating at the cranial end of a piece of gut and passing caudally. In man the peristaltic wave always travels in the same direction. The function of the peristaltic wave is to move the contents of the gut along the tube in the direction of the wave. In the oesophagus peristaltic waves are initiated by swallowing. Gases in the stomach may result from the action of the digestive secretions on foodstuffs or may even reach the stomach from subsequent parts of the digestive tract. Most commonly, they are simply air that has been swallowed with the food. Such gases may be eliminated if they become uncomfortable by eructation.
Anatomy
The digestive system, or the gut, is a muscular tube lined with endoderm which traverses the body from mouth to anus, sealed at various points by muscular sphincters controlled by the autonomic nervous system. The cephalic end of the tube, the mouth cavity, is closed by the lips; entry to the tube is therefore under voluntary control. The caudal end, the anus, is sealed by the anal sphincters, the outermost of which is under voluntary control in the adult. Since the tube may be open to the external environment at both ends, its inside and contents must be regarded as external to the body. Food and drink are taken into this tube, where they are physically and chemically processed. Their components are then absorbed actively or passively across its wall into the body cavity proper, or else remain unabsorbed and are carried down the tube to be excreted as faeces. The walls of the gut are made up of muscle whose contractions knead and mix the food after the initial chopping and grinding in the mouth, and also propel the food mixture onwards. The mural epithelium is specialised to form glands of varying complexity: these produce mucoid substances to lubricate the passage of the food mixture and to protect the walls, and enzymes which break down complex foodstuffs into simple molecules. The glands may be situated in the thickness of the walls or may be so deeply invaginated as to form separate structures – as, for example, the pancreas. Other parts of the tube are specialised for absorption, with infoldings of the surface mucous membrane (the mucosa) which enormously increase the area available for diffusion. The mucosa, like other epithelial surfaces, is constantly losing its outer layer of cells and replacing them. This helps to protect it against the abrasion due to the passage of food. The cells lost in this way in the lower part of the gut are excreted in the faeces, together with some of the gut bacteria and any secretions which have not been reabsorbed.
The digestive tube is subdivided into a number of organs on a basis of the anatomical and functional breaks due to the sphincters (Fig. 11.1). The mouth, or oral cavity, is separated from the oro-pharynx only by a ring of lymphoid tissue and is continuous through the naso-pharynx with the nasal cavity. The pharynx is not anatomically separated from the oesophagus, although a functional sphincter, the cricopharyngeal sphincter, has been described. The oesophagus extends from there to the cardiac sphincter at the entrance to the stomach. It is situated within the thoracic cavity and at rest its lumen is in equilibrium with thoracic pressures because of the thinness of its walls. A balloon in the oesophagus may therefore be used as a pressure transducer to measure intrathoracic pressures. The next portion is the stomach, separated above from the oesophagus by the cardiac, or gastroesophageal sphincter, and below from the small intestine by the gastroduodenal, or pyloric, sphincter. Functionally the stomach has a cardiac portion, a fundus or body, and the tubular pyloric antrum leading to the pylorus. The small intestine is made up of the short C-shaped duodenum holding the pancreas within its loop, and the longer jejunum and ileum. Since there is no obvious division between jejunum and ileum their lengths are arbitrarily estimated as in the ratio of 2:3. The small intestine as a whole is about 270 cm long in the living subject. Its termination is marked by the ileocaecal valve or sphincter. The ileum projects into the colon (the next part of the gut) and since it enters laterally, the opening is closed by colonic contractions. The colon extends a short distance downwards as the caecum, which ends in a small blind-ended extension, the appendix. The colon proper passes up towards the liver, crosses the abdominal cavity from right to left and then continues downwards to join the rectum at the colonic sphincter. The rectum is the final part of the gut and ends in the anus with the internal anal sphincter.
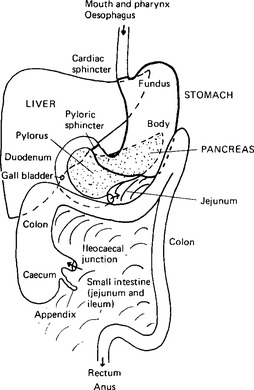
Figure 11.1 The main organs of the digestive system as seen when the abdomen is opened. The outlines are continued as dotted lines when they pass behind another organ: the complex folds of the small intestine are indicated by curved lines of shading, and only the beginning of the jejunum and the end of the ileum, where it joins the caecum, are shown for localisation.
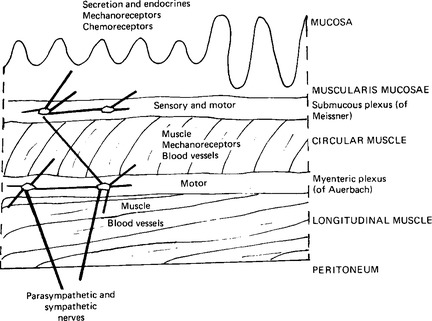
The major part of the tube from oesophagus to anus is made up of layers of muscle separated from each other by connective tissue in which lie blood vessels and nerves. Characteristically the muscle is arranged in three layers: the submucosal, the circular and the longitudinal layers (Fig. 11.2). In fact, both circular and longitudinal layers are spiral muscles, the circular a tightly coiled spiral, and the longitudinal an extended spiral. The lumen of the gut is lined with a mucosa whose surface varies from a stratified squamous epithelium in the mouth and oesophagus to a columnar epithelium in the intestine. The lamina propria is invaginated by glands producing mucus and digestive secretions. The blood vessels and lymphatics are specially arranged in the parts of the intestine whose function is primarily absorptive. Although the gut is composed of smooth muscle with its own intrinsic rhythmicity, it has an extensive innervation to modify and co-ordinate its activity as well as to control its secretions. Two nerve plexuses are described: a submucous plexus (of Meissner) between the muscularis mucosae and the circular muscle layer; and the myenteric plexus (of Auerbach) between the circular and longitudinal muscle layers. These plexuses are both sensory and motor. The extrinsic nerves connect mainly with the myenteric plexus. Parasympathetic fibres travel in the vagus to the upper part of the gut, and in the pelvic and splanchnic nerves to the colon and rectum (Fig. 9.18). Sympathetic fibres pass from the coeliac plexus to the stomach and small intestine, from the superior mesenteric plexus to the small intestine, caecum, ascending and transverse colon, and from the inferior mesenteric plexus to the descending colon and the rectum (Fig. 9.19).
Movements of the Digestive Tract
The control and co-ordination of the processes of ingestion, digestion and excretion are discussed later (Chaps. 19 and 21) but a general description of the muscular activities seen in the gut is given here.
The movements of the muscularis mucosae are independent of those of the longitudinal and circular muscles: they originate as responses to the contact of the mucosal surface with any material within the gut lumen. Extrinsic nerves, however, are able to modify the excitability of the muscularis mucosae via the nerve plexuses in the same way as they modify the excitability of the longitudinal and circular muscles: the parasympathetic generally increasing, and the sympathetic inhibiting, activity.
The stomach in the resting state is of small volume – perhaps 50ml – but as it fills it stretches, showing the plastic and elastic properties of smooth muscle (see Chap. 5). As food passes into the stomach, layering occurs in the fundus, but by the time the food reaches the pylorus mixing has taken place. The appearance of the stomach in X-ray pictures depends upon the density of its contents – the stomach as a whole is equivalent to a bag of fluid in a fluid-filled cavity and if its density is low it floats. Within the stomach the same principle holds good: solid materials sink within the fundus, liquids ‘float’, and gases rise to the top – to give the appearance of a cap in some X-ray pictures. The passage of solid material, however, towards the pyloric antrum is relatively slow. Although the rate of emptying varies, when it begins each peristaltic wave transfers about 1ml to the duodenum, resulting in a transfer rate of about 20 ml/min. About 35% of a 500 ml fluid meal leaves the stomach in one hour and it usually takes around 4h for a normal meal to leave the stomach. Patients about to undergo general anaesthesia are therefore advised to refrain from meals for some 4h before the operation, so that the stomach may be empty and the risk of vomiting (with the consequent danger of vomitus passing into the respiratory tract) reduced.
Secretions of the digestive tract – the salivary glands and saliva
The secretions of the oral cavity are collectively known as whole or mixed saliva, although some writers prefer the term ‘oral fluid’. All three terms refer to the fluid which is collected by expectoration; secretions of individual glands are named appropriately as parotid saliva, accessory gland saliva, etc. In the resting mouth the lips and cheeks are almost in contact with the teeth anteriorly, and the tongue is in close proximity with the teeth and the palate posteriorly. Only a thin film of whole saliva separates these structures and this constitutes their immediate environment. How far saliva can be considered the environment of the teeth themselves is debatable, since most of the dental enamel surfaces are covered either by a glycoprotein pellicle or the bacterial film known as dental plaque. However, water and small molecules appear to diffuse easily through thin layers of dental plaque.
Mixed saliva, whole saliva or oral fluid
Components of the oral fluid
Whole saliva is made up of the secretions of the three pairs of major salivary glands – the parotid, the submandibular and the sublingual – together with the secretions of the minor, or accessory, glands which are distributed in the mucosa of the cheek, lips, hard and soft palates, and tongue. In addition to these secretions, the total oral fluid includes the gingival crevicular fluid, which passes between the teeth and the gingival cuff tissue to reach the oral cavity (p. 23), and, if it is formed by human teeth, enamel fluid (p. 23).
The salivary glands
Anatomy
Each parotid gland is pyramidal in shape with the base of the pyramid lying directly below the skin (Fig. 11.3). Each is enclosed in a fibrous capsule and weighs about 25 g. The main duct passes forward over the masseter muscle, turns inward to pierce the buccinator muscle and then terminates in a small papilla on the mucosa of the cheek close to the upper first molar tooth. The situation of the duct renders it easy to attach a cup (a Lashley or Carlsen-Crittenden cup) in position over it by suction.
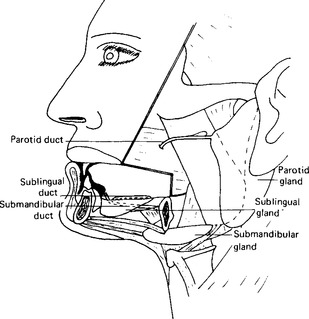
The submandibular glands, irregular in shape, are described as being of the size of walnuts. They lie posteriorly in the floor of the mouth (Fig. 11.3
Stay updated, free dental videos. Join our Telegram channel
VIDEdental - Online dental courses


