Development of teeth
Learning objectives
■ Describe the origin of the tooth formative cells and the role of induction in tooth formation.
■ Describe the stages of tooth formation and the mineralization of enamel and dentin.
■ Describe the development of the tissues that surround the developing teeth.
Key terms
Alveolar bone proper
Ameloblasts
Amelogenin
Bell stage
Bud stage
Cap stage
Cell signaling
Cementoid
Dental follicle
Dental lamina
Dental papilla
Dental pulp
Dentin
Desmosomes
Developmental or primary cuticle
Enamelin
Epithelial diaphragm
Epithelial rests
Epithelial root sheath/Hertwig’s epithelial root sheath (HERS)
Hemidesmosomes
Inner enamel epithelial cells
Intermediate cementum
Interradicular bone
Neural crest cells
Odontoblast process
Odontoblasts
Outer enamel epithelial cells
Predentin
Protective stage
Pulp proliferation zone
Reduced enamel epithelium
Stellate reticulum cells
Stratum intermedium cells
Successional lamina
Supporting bone
Terminal bar apparatus
Tomes processes
Overview
The human is a species with two sets of dentition (diphyodont) that normally include 20 primary and 32 permanent teeth that develop from the interaction of the oral ectodermal cells and the underlying mesenchymal cells. Each developing tooth grows as an anatomically distinct unit, but the basic developmental process is similar for all teeth.
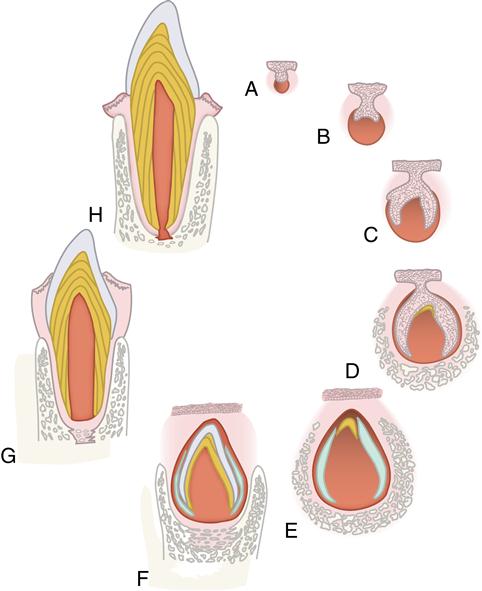
Each tooth develops through successive bud, cap, and bell stages (Fig. 5-1, A to C). During these early stages, the tooth germs grow and expand, and the cells that are to form the hard tissues of the teeth differentiate. Differentiation takes place in the bell stage, setting the stage for enamel and dentin formation (Fig. 5-1, D and E). As the crowns are formed and mineralized, the roots of the teeth begin to form. After the roots calcify, the supporting tissues of the teeth—the cementum, periodontal ligament, and alveolar bone—begin to develop (Fig. 5-1, F and G). This formation occurs whether the tooth is an incisor with a single root, a premolar with several roots, or a molar with multiple roots. Subsequently, the completed tooth crown erupts into the oral cavity (Fig. 5-1, G). Root formation and cementogenesis continue until a functional tooth and its supporting structures are fully developed (Fig. 5-1, G and H).
 CLINICAL COMMENT
CLINICAL COMMENT
Developmentally missing permanent teeth can be a result of a genetic abnormality. When fewer than six teeth are missing, it is termed hypodontia, and when more than six teeth are missing, it is called oligodontia.
Initiation of tooth development
Teeth develop from two types of cells: oral ectodermal cells form the enamel organ and mesenchymal cells form the dental papilla. Enamel develops from the enamel organ, and dentin forms from the dental papilla. The interaction of these epithelial and mesenchymal cells is vital to the initiation and formation of the teeth. In addition to these cells, the neural crest cells contribute to tooth development. The neural crest cells arise and begin their migration from the mesencephalic portion of the developing neural tube at an early stage of development and migrate into the jaws, intermingling with mesenchymal cells. They function by integrating with the dental papillae and epithelial cells of the early enamel organ, which aids in the development of the teeth. The cells also function in the development of the salivary glands, bone, cartilage, nerves, and muscles of the face. Chapter 1 discussed neural crest cells and explained the cell’s migration (see Fig. 1-17).
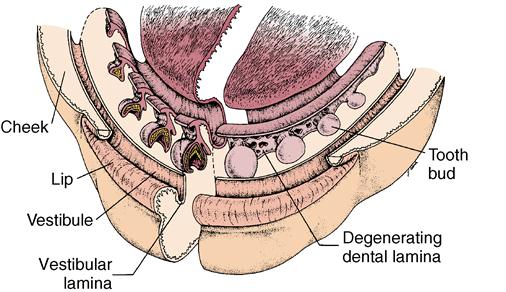
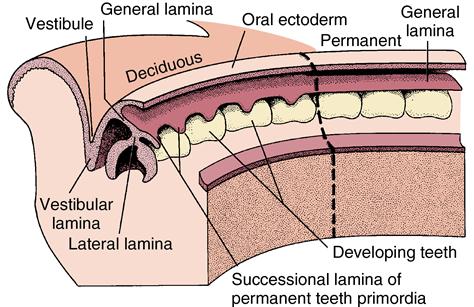
The first sign of tooth formation is the proliferation of ectodermal cells overlying specific areas of the oral ectoderm; their proliferation results in the development of dental lamina. The dental lamina develops into a sheet of epithelial cells that pushes into the underlying mesenchyme around the perimeter of both the maxillary and mandibular jaws beginning at the midline (Fig. 5-2). At the leading edge of the lamina, 20 areas of enlargement appear, which form tooth buds for the 20 primary teeth (see Fig. 5-2). At this early stage, the tooth buds have already determined their crown morphology of an incisor or molar. This is the result of a complicated series of gene expression that alternates between the epithelium and the mesenchymal tissue. After primary teeth develop from the buds, the leading edge of the lamina continues to grow to develop the permanent teeth, which succeed the 20 primary teeth. This part of the lamina is thus called the successional lamina (Fig. 5-3). The lamina continues posteriorly into the elongating jaw, and from it come the posterior teeth, which form behind the primary teeth. In this manner, 20 of the permanent teeth replace the 20 primary teeth, and 12 posterior permanent molars develop behind the primary dentition (see Fig. 5-3). The last teeth to develop are the third molars, which develop about 15 years after birth. Because the permanent molars do not succeed the primary teeth, they do not form from the successional lamina but from the general lamina and are termed accessional teeth. The initiating dental lamina that forms both the successional and general lamina begins to function in the sixth prenatal week and continues to function until the fifteenth year, producing all 52 teeth. In general, the teeth develop anteroposteriorly, which relates to the growing jaws. The posterior molars do not develop until space is available for them in the posterior jaw area. The second dentition does not develop until after the primary teeth are formed and functioning. Gradually the permanent teeth form under the primary crowns and later posteriorly to the primary molars.
Stages of tooth development
Although tooth formation is a continuous process, it is characterized by a series of easily distinguishable stages known as the bud, cap, and bell stages. Each stage is defined according to the shape of the ectodermally derived enamel organ, which is a part of the developing tooth. The initial stage, the bud stage, is a rounded, localized growth of ectodermal cells surrounded by proliferating mesenchymal cells (Fig. 5-4). Gradually, as the rounded epithelial bud enlarges, it gains a concave surface, which begins the cap stage (Fig. 5-5). The ectodermal then differentiate to become the enamel organ and remain attached to the lamina. The mesenchyme forms the dental papilla, which becomes the dental pulp. The tissue surrounding these two structures is the dental follicle.
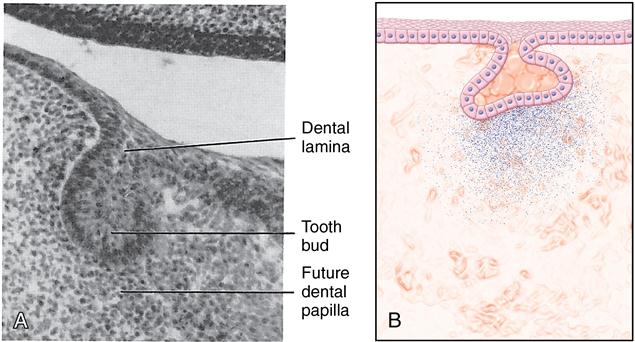
A, Histology of the bud stage. B, Diagram of the bud stage. (B from Hargreaves KM, Cohen S: Cohen’s pathways of the pulp, ed 10, St. Louis, 2011, Mosby.)
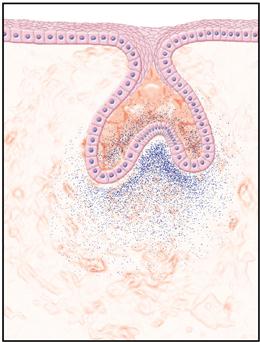
The enamel organ is outlined in blue, indicating that it is of ectodermal origin. The mesenchyme of the dental papilla surrounds the enamel organ. (From Hargreaves KM, Cohen S: Cohen’s pathways of the pulp, ed 10, St. Louis, 2011, Mosby.)
After further growth of the papilla and the enamel organ, the tooth reaches the morphodifferentiation and histodifferentiation stage, also known as the bell stage (Fig. 5-6). At this stage, the inner enamel epithelial cells are characterized by the shape of the tooth they form (see Fig. 5-6). Also, the cells of the enamel organ have differentiated into the outer enamel epithelial cells, which cover the enamel organ, and inner enamel epithelial cells, which become the ameloblasts that form the enamel of the tooth crown. Between these two cell layers are the stellate reticulum cells, which are star shaped with processes attached to each other. A fourth layer in the enamel organ is composed of stratum intermedium cells. These cells lie adjacent to the inner enamel epithelial cells. They assist the ameloblast in the formation of enamel. The function of the outer enamel epithelial cells is to organize a network of capillaries that will bring nutrition to the ameloblasts.
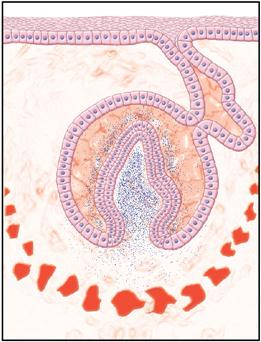
During this stage of tooth development, both odontoblasts and ameloblasts have fully differentiated in the cuspal region(s). Blood vessels develop in the dental papilla, whereas the only vascularized structure in the enamel organ is the outer enamel epithelium that contains a capillary plexus. (From Hargreaves KM, Cohen S: Cohen’s pathways of the pulp, ed 10, St. Louis, 2011, Mosby.)
From the outer enamel epithelium, nutrients will percolate through the stellate reticulum to the ameloblasts. During the bell stage, cells in the periphery of the dental papilla become odontoblasts. These cells differentiate from neural crest cells or synonymously, ectomesenchymal cells. After the odontoblasts elongate, become columnar and polarize, they form an extracellular matrix (ECM) of 90% type I collagen fibers and 10% noncollagenous proteins termed predentin. After 24 hours, this increment of matrix calcifies and becomes dentin. When several increments of dentin have formed, the differentiated ameloblasts deposit an enamel matrix. Dentinogenesis always precedes amelogenesis. After the enamel organ is differentiated, the dental lamina begins to degenerate by undergoing autolysis. The dental lamina disappears in the anterior part of the mouth, although it remains active in the posterior region for many years (see Figs. 5-2 and 5-3).
Cells interact through a system of effectors, modulators, and receptors called cell signaling. An example of such a system is epithelial–mesenchymal interaction in tooth development. The precursor cells, odontoblast and ameloblast, establish a positional relationship by means of effectors and receptors that are on the cell surface. The ameloblast differentiates first, causing the precursor odontoblast to locate itself adjacent. Then the odontoblast differentiates, establishing with the ameloblast a basement membrane that then forms a dentinal matrix. After this formation occurs, the ameloblast forms the enamel matrix. Thus it is not only cells, but also basal lamina and dentin matrix, that contain substances that cause cell changes and position.
Development of the dental papilla
Densely packed cells characterize the dental papilla. This is evident even in the early bud stage, during which cells proliferate around the enlarging tooth buds at the leading edge of the dental lamina (Fig. 5-7). The papilla cells are believed to be significant in furthering enamel organ bud formation into the cap and bell stages. This cell density is maintained as the enamel organ grows. Cells of the dental papilla are found on close examination to be fibroblasts and appear to be in a delicate reticulum (Fig. 5-8). Blood vessels appear early in the dental papilla, initially in the central region along with postganglion sympathetic nerve fibers associated with these vessels. The vessels bring nutrition to the rapidly growing organ. As the papilla grows, smaller vessels are also seen in the periphery of the organ, bringing nutrition to the elongating odontoblasts (see Fig. 5-6). Cellular changes result in formation of a mineralized tissue around the central papilla. As this occurs, the papilla becomes known as the dental pulp.
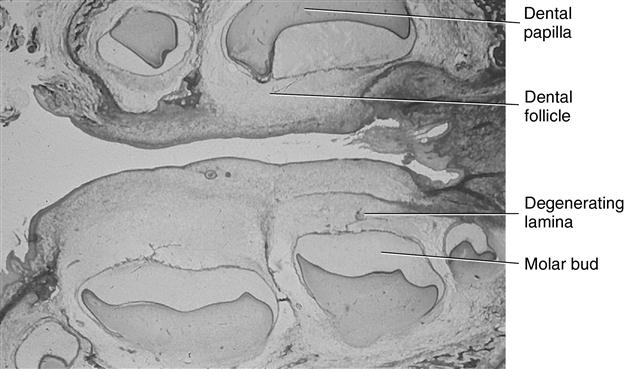
Sagittal view of the human maxillary and mandibular molar tooth buds.
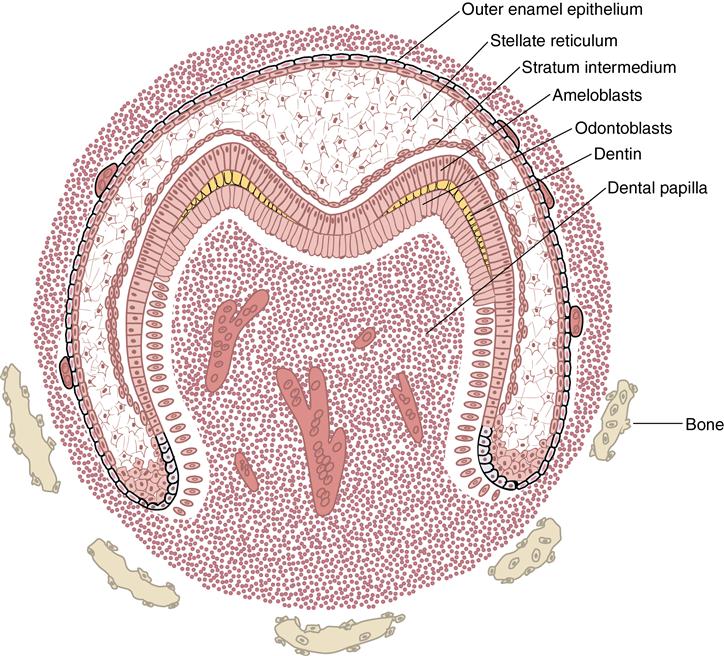
The initial formation of dentin (yellow) at the cuspal tips and the vascularized pulp organ are characteristic of the dentinogenesis stage. The dental follicular cells are differentiating around the enamel organ and alveolar bone proper is beginning to define the dental crypt.
Dentinogenesis
As the odontoblasts elongate, they gain the appearance of a protein-producing cell. A process develops at the proximal end of the cell, adjacent to the dentinoenamel junction. Gradually the cell moves pulpward, and the cell process, known as the odontoblast process, elongates (Fig. 5-9). The odontoblast becomes active in dentinal matrix formation similar to an osteoblast when it moves away from a spicule of bone. Increments of dentin are formed along the dentinoenamel junction. The dentinal matrix is first a meshwork of collagen fibers, but within 24 hours it becomes calcified. It is called
Stay updated, free dental videos. Join our Telegram channel
VIDEdental - Online dental courses


