Chapter 4 The Abdomen, Pelvis, and Perineum
The Abdomen, Pelvis, and Perineum
1 Skeleton and Subdivisions
SKELETON OF THE ABDOMEN
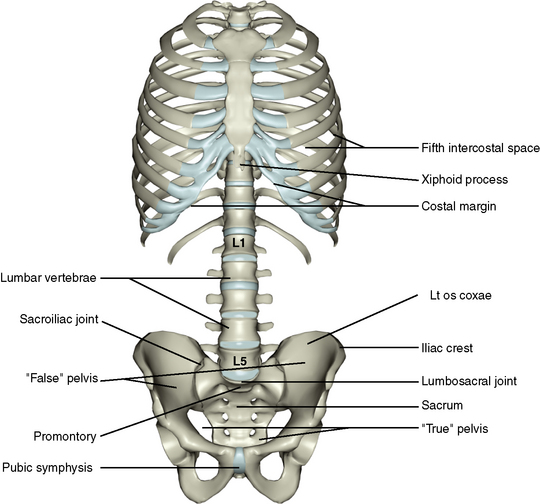
The skeleton of the abdomen consists of a thoracic component, a vertebral component, and a pelvic component (Figure 4-1).
Thoracic Component
Because the abdominal cavity extends up into the thoracic cage, the thorax between the fifth intercostal space above to the costal margin below should be considered, along with the abdominal skeleton. The thoracic cage is discussed on pages 54 to 57.
Vertebral Component
The lower thoracic vertebrae, the lumbar vertebrae, the sacrum, and the coccyx form the vertebral component of the abdominal skeleton. These structures are described on pages 40 to 43.
Pelvic Component
Os Coxae (Hip Bone)
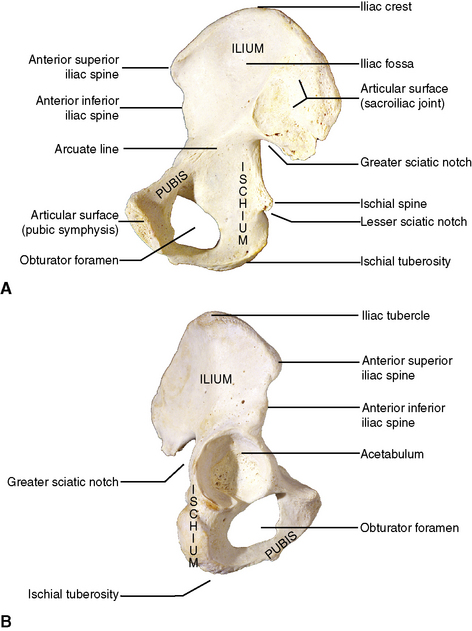
The right and left os coxae comprise the lower limb girdle and take part in the hip joint (see Chapter 10). In addition, they join with the sacrum and the coccyx to form the pelvic cavity, which houses and protects several pelvic viscera. The os coxae is originally formed from three separate bones that fuse as one complete bone by about the sixteenth year. The bones are the ilium, ischium, and pubis, and they lend their names to the three regions of the single adult os coxae (Figure 4-2).
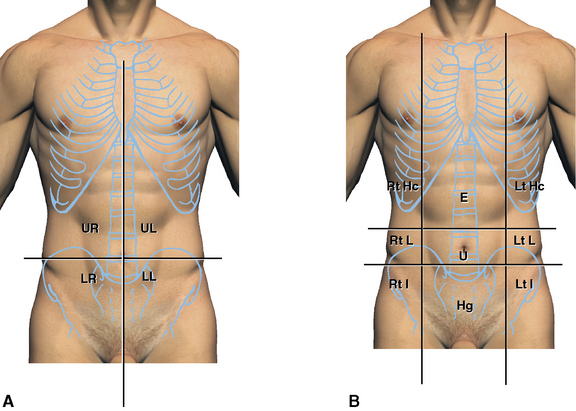
DIVISIONS OF THE ABDOMEN
To facilitate descriptions of abdominal visceral locations, the abdomen is divided into regions (Figure 4-3). There are two systems in common use. One divides the abdomen into four quadrants based on the median sagittal plane in the abdomen intersecting with a transverse plane. This effectively divides the abdomen into upper left, upper right, lower left, and lower right quadrants.
2 The Abdominal Walls
ANTEROLATERAL ABDOMINAL WALL
Surface Features
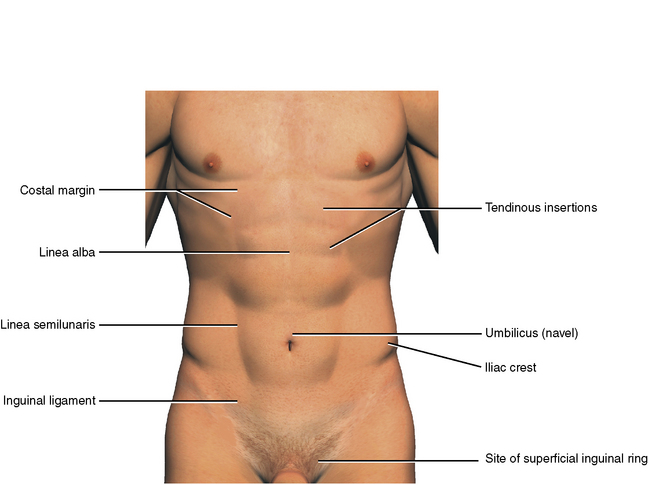
Palpable abdominal landmarks are the costal margins and the xiphoid process, the iliac crests, the superior and inferior anterior iliac spines, and the pubic tubercles (Figure 4-4). In thin, muscular individuals a linear depression runs in the midline of the abdomen from the xiphoid process to the symphysis pubis. It represents the underlying linea alba, which is the union of right and left muscular aponeuroses. The linea semilunaris is a curved line lateral to the midline. It represents the lateral limits of the rectus abdominis muscle. Transverse bands running from the linea semilunaris and the midline represent underlying tendinous insertions of the rectus abdominis muscle. The umbilicus (belly button) is the scarred result of the postnatal closure of the umbilical cord. A slight crease runs from the anterior superior iliac spine toward the pubic tubercle. It represents the position of the inguinal ligament. Just above and medial to the pubic tubercle is the site of the superficial inguinal ring, the site of indirect inguinal hernias.
Layers
From outward within, the layers that compose the anterolateral abdominal wall below the skin are the (1) superficial fascia, (2) deep fascia, (3) muscles and aponeurosis, (4) transversalis fascia, (5) extraperitoneal layer, and (6) peritoneum (Figure 4-5).
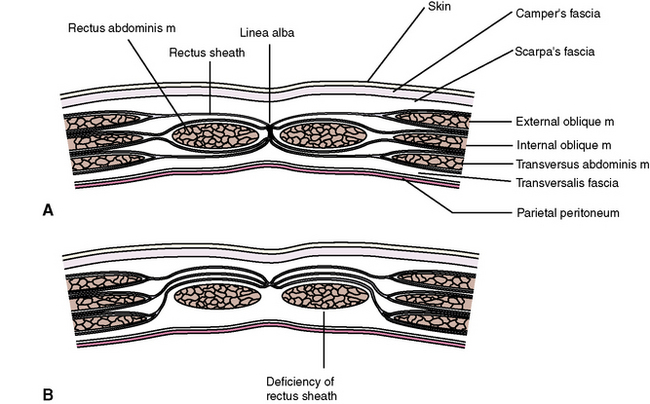
Deep Fascia
The deep fascia of the abdomen is even more membranous and covers the underlying muscle layer.
Muscles and Aponeuroses
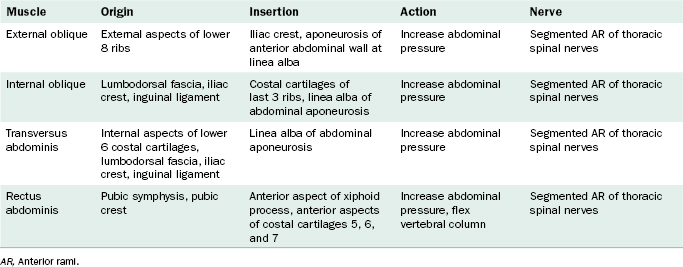
Four pairs of bilateral muscles and their flattened tendons, or aponeuroses, contribute to the anterolateral abdominal wall (Table 4-1 and Figure 4-6). There are three pairs of flat muscles (external oblique muscle, internal oblique muscle, and transverse abdominis muscle) and one pair of straplike muscles (rectus abdominis muscle). The three flat muscles are layered in sheets, but the fiber directions of each muscle run in different directions, resulting in a strong laminated muscular unit. Each muscle is fleshy laterally and forms membranous aponeuroses medially. As the aponeuroses of the three flat muscles approach the midline, they form a membranous sheath that wraps around the rectus abdominis muscle.
The formation of the rectus sheath is rather complicated, as shown in Figure 4-5. Two representative cuts through the muscle layer are shown. At level A the internal oblique apilioneurosis splits to encircle the rectus abdominis muscle. At level B all of the aponeuroses are anterior to the muscle, leaving a deficiency in the sheath posteriorly. The arcuate line marks the limit of the aponeurotic contribution to the posterior wall of the bed of the rectus sheath. Below this arcuate line, only the transversalis fascia separates the rectus abdominis muscle from the underlying peritoneum.
Peritoneum
The deep, fatty, extraperitoneal layer beco-mes membranous and continuous with the more organized connective tissue of the peritoneal layer below. Deeper still is a smooth, glistening layer of mesothelial cells, which line the peritoneal cavity. Peritoneum and the contents of the abdomen are discussed on page 99.
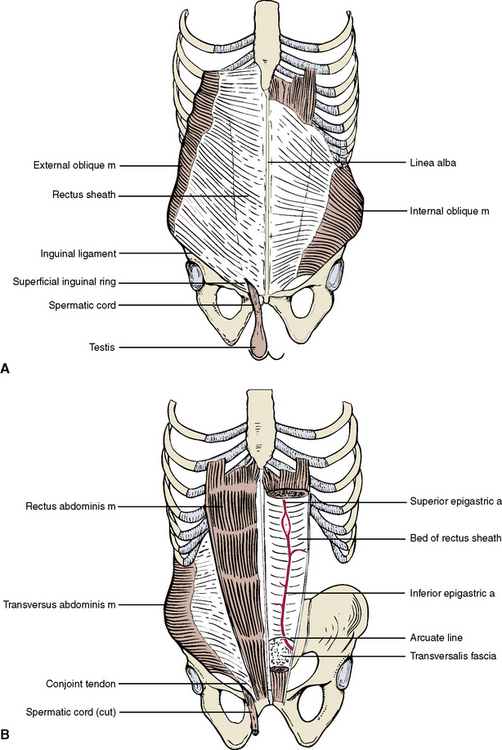
Inguinal Region
The inguinal region is superior to the medial portion of the inguinal ligament. Here the lower fibers of the internal oblique and transversus abdominis muscles do not insert into the inguinal ligament but rather insert as the conjoint tendon into the pubis, leaving a free inferior gap (see Figure 4-6). The external oblique aponeurosis covers this gap incompletely, its inferior medial fibers diverging as a triangular opening as it approaches the pubis. The opening is the superficial inguinal ring, which serves as the external opening of the inguinal canal. The canal is formed by the deficiency of the transversus abdominis and internal oblique muscles and their conjoint tendon posteriorly and the external oblique aponeurosis anteriorly. Issuing from this canal in males is the spermatic cord, from which the testis is suspended within the scrotum. In females the round ligament of the uterus passes through the canal to attach to the labium majus.
Spermatic Cord
The gonads develop within the extraperitoneal layer of the abdomen (Figure 4-7). The gubernaculum extends from this site down to the developing scrotum in males and to the labia majora in females. It marks the path for the descent of the testes in males.
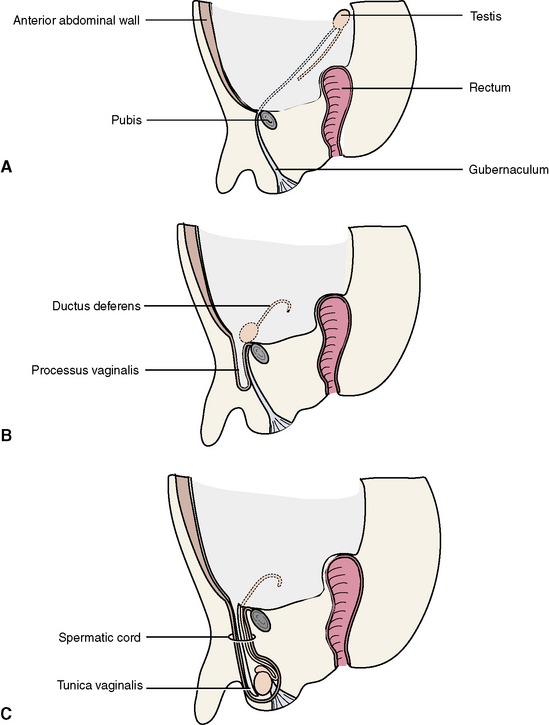
Layers
Blood and Nerve Supply
The arterial supply is from two sources. Posteriorly segmented branches of the aorta follow the spinal nerves. Anteriorly segmented branches arise from the superior and inferior epigastric arteries running in the bed of the rectus sheath (see Figure 4-6). The superior epigastric artery arises superiorly as a continuation of the internal thoracic artery; the inferior epigastric artery arises inferiorly from the external iliac artery. The two epigastric arteries anastomose above the umbilicus and supply the contents of the rectus sheath and the sheath itself. Collateral branches from the epigastric arteries anastomose with the aortic collateral vessels posteriorly.
SUPERIOR ABDOMINAL WALL
Origins of the Thoracic Diaphragm
The muscular slips of the diaphragm originate from three sites of attachment (Figure 4-8). Sternal slips arise from the posterior aspect of the xiphoid process. Costal slips originate from the internal surfaces of the lower six costal cartilages and the twelfth rib. Lumbar attachments arise as a right crus from the vertebral bodies and discs of L1, L2, and L3, and a left crus arising from the vertebral bodies and discs of L1 and L2. The two crura cross each other to form the median arcuate ligament through which enters the abdominal aorta. Tendons arching from the crura to the transverse processes of L1 form medial arcuate ligaments. Tendons arching from the transverse processes of L1 to the midpoints of the twelfth ribs form lateral arcuate ligaments.
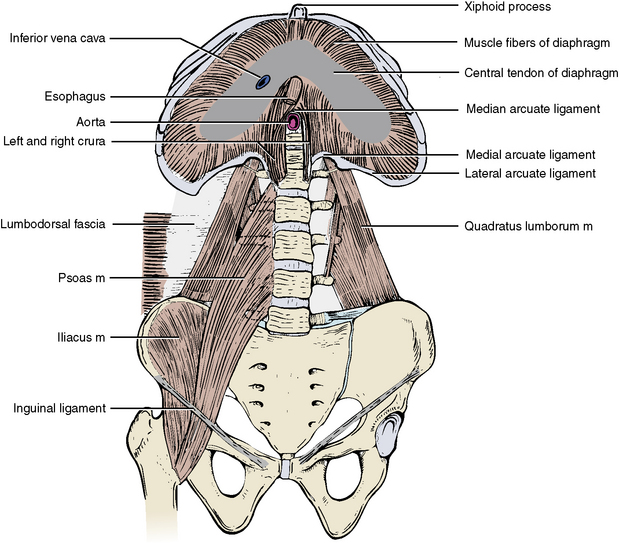
Structures Passing Through the Diaphragm
Functions of the Diaphragm
The main function of the diaphragm is respiration. The contracting muscle fibers pull down on the central tendon during thoracic inspiration. The mechanisms involved with inspiration and expiration are discussed in Chapter 3, page 67.
Nerve Supply of the Diaphragm
The diaphragm originates early in development from the cervical region, and as it descends during development it drags its cervical nerve and blood supply along with it. The phrenic nerve is the motor and sensory supply to the diaphragm. It arises in the neck from anterior rami of spinal nerves C3, C4, and C5. The phrenic nerve descends through the thoracic inlet and travels inferiorly on either side of the middle mediastinum to reach the diaphragm (see Chapter 3, page 84).
POSTERIOR ABDOMINAL WALL
The posterior abdominal wall consists of skin and fascia, bone (including the lumbar and thoracic vertebrae, the lower ribs, and the os coxae), and a fleshy component consisting of three muscles (see Figure 4-8).
The psoas major and iliacus muscles are generally considered as a unit (iliopsoas muscle), and it is actually a muscle of the lower limb described in Chapter 10.
INFERIOR ABDOMINAL WALL
Muscles
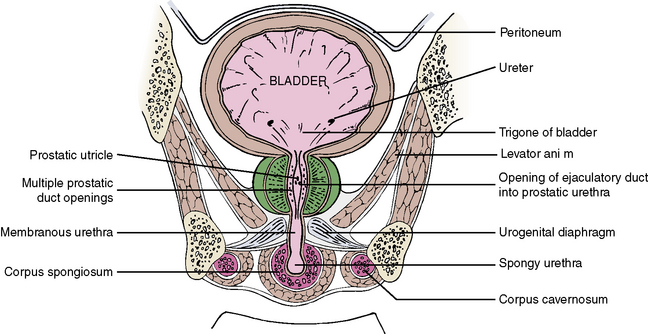
The levator ani muscles originate along the internal aspects of the os coxae (Figure 4-9 and Table 4-2). The fibers run medially and inferiorly toward the rectum to blend with the longitudinal smooth muscle of the rectum. A portion of each levator ani muscle runs from the ischial tuberosity to the coccyx and is given a special name, the coccygeus muscle.
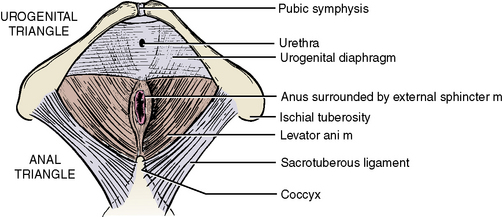
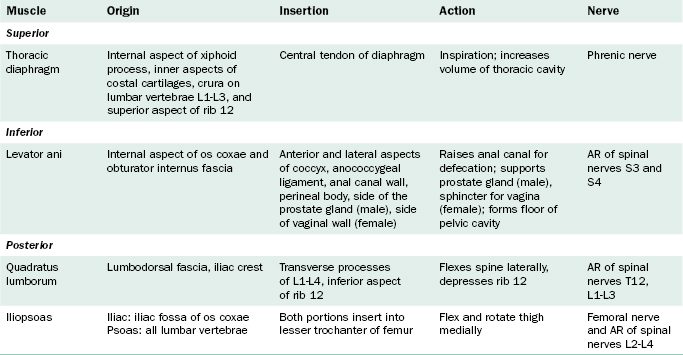
The levator ani muscles help draw the rectum superiorly during defecation. The muscle is deficient anteriorly to allow passage of the urethra in men and the urethra and the vagina in women (see also Figure 4-37).
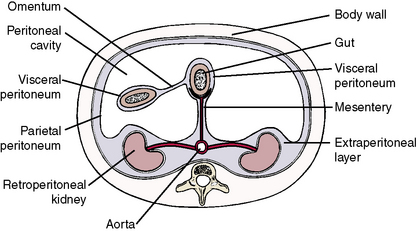
3 Peritoneum and the Peritoneal Cavity
NOMENCLATURE
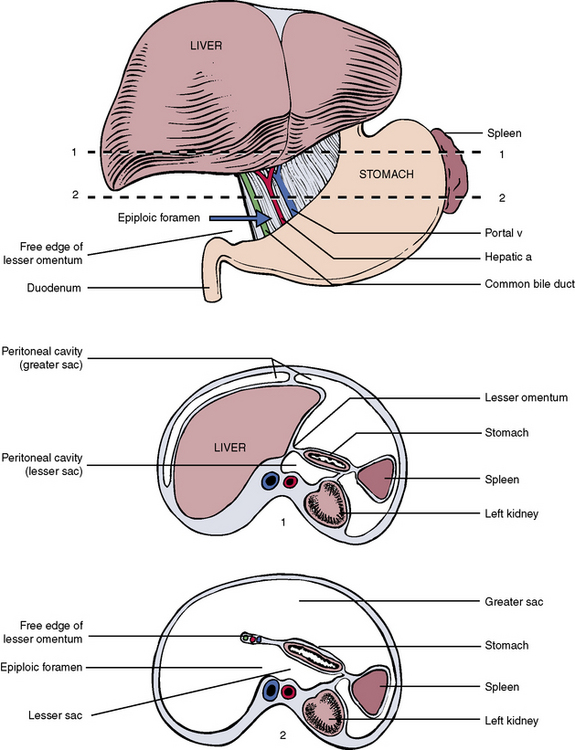
To understand the distribution of peritoneum within the hollow abdominal cavity, consider the schematic representation of a transverse section through the abdomen in Figure 4-10.
DEVELOPMENT OF THE GUT AND PERITONEUM
To better understand the complexities of the peritoneal linings, it is advantageous to consider the development of the gut during embryonic life. Figure 4-11 shows the primitive gut, which is essentially a tube suspended from the posterior body wall by a dorsal mesentery. Running to the suspended gut through the dorsal mesentery are three branches of the abdominal aorta: (1) the celiac trunk, which supplies the foregut; (2) the superior mesenteric artery, which supplies the midgut; and (3) the inferior mesenteric artery, which supplies the hindgut.
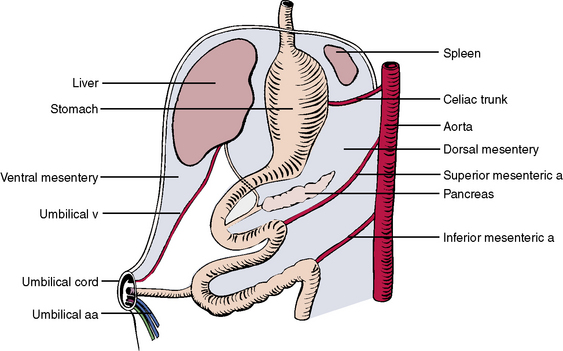
Figure 4-12, A, shows a transverse section through the developing abdomen. As indicated, the stomach and spleen move to the left, and the liver moves to the right. Figure 4-12, B, represents a later stage in development, with the liver now occupying most of the upper right quadrant and the stomach and spleen occupying the upper left quadrant. In addition, the mesentery of the stomach becomes adherent to the posterior body wall and is lost. In like manner, the mesenteries of the duodenum, ascending colon, descending colon, and rectum are fused to the posterior body wall, and these sections of gut become “retroperitoneal.” With blunt dissection, these obliterated mesenteries can be restored in the laboratory and occasionally in actual abdominal surgery (as needed).
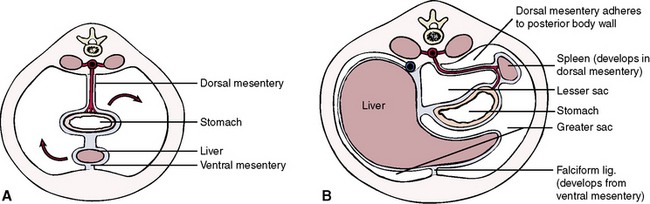
The liver does not have a mesentery, nor is it considered retroperitoneal. Rather, it is suspended from the diaphragm and posterior body wall by a broad-based peritoneal attachment, called the coronary ligament, which is described along with the liver (on page 115).
4 Blood and Nerve Supply to the Abdomen
ARTERIAL SUPPLY: THE ABDOMINAL AORTA
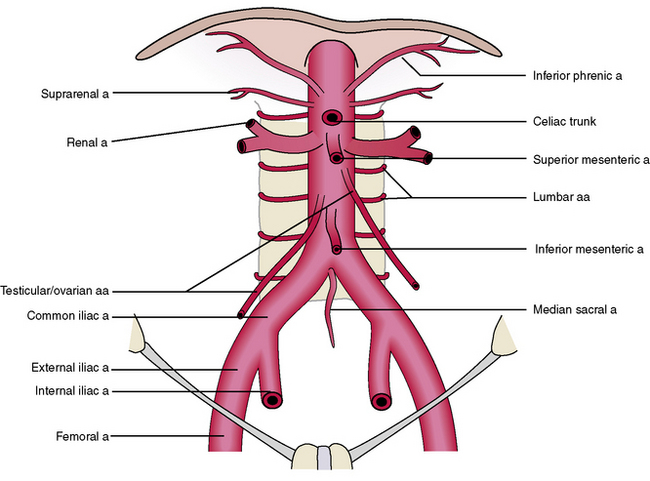
The descending aorta of the thorax passes through the diaphragm to become the abdominal aorta (Figure 4-13). It descends on the posterior body wall to the pelvis, and at vertebral level L4 it divides into two terminal branches: the right and left common iliac arteries. These, in turn, divide into the right and left external iliac arteries, which descend to supply the lower limb, and the right and left internal iliac arteries, which supply pelvic structures. The abdominal aorta gives off a number of branches within the abdomen before terminating as the common iliac arteries at vertebral level L4.
VENOUS RETURN: THE INFERIOR VENA CAVA
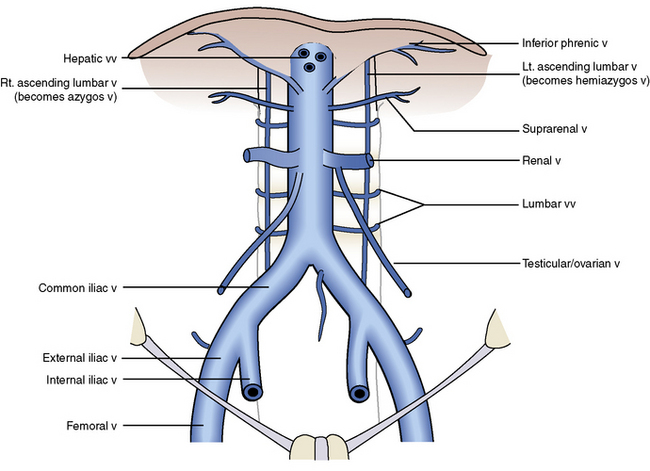
The external iliac veins, which drain the lower limbs, and the internal iliac veins, which drain the pelvis, unite within the pelvis to form the right and left common iliac veins (Figure 4-14).
Somatic Branches
The somatic branches correspond to the somatic arterial branches.
Tributaries of the Gastrointestinal Tract and Associated Glands
Venous return from the gastrointestinal tract does not return directly to the inferior vena cava (see Figure 4-27). Instead, veins returning from the gut join to form the portal vein. The portal vein enters the liver, carrying nutrients absorbed from the gut. Within the liver the portal vein ultimately ends as a capillary bed; at this level, nutrients are exchanged for processing and storage within the liver. The portal capillary beds are drained by hepatic veins, which leave the liver to enter the inferior vena cava as several hepatic veins.
NERVES OF THE ABDOMEN
Somatic Nerves
The anterior rami of the lower six thoracic spinal nerves and the first lumbar nerve supply the various layers of the anterolateral abdominal walls. Two branches of the anterior rami of L1, the iliohypogastric and ilioinguinal nerves, supply the lower portion of the abdominal wall (see Figure 10-20).
The anterior rami of lumbar nerves L1 to L4 unite and divide within the substance of the psoas muscle to form the lumbar plexus. Branches of the lumbar plexus supply a portion of the lower limb and consequently are considered in Chapter 10 with the lower limb.
Stay updated, free dental videos. Join our Telegram channel
VIDEdental - Online dental courses


