Chapter 1 General Concepts
General Concepts
1 The Study of Anatomy
2 Terminology
THE ANATOMICAL POSITION
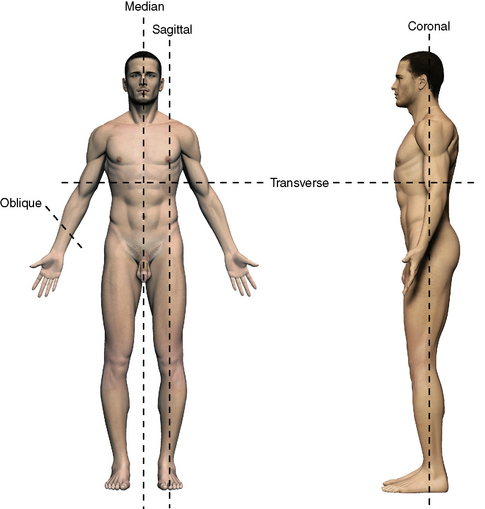
In the dissecting laboratory, we assume, by convention, that our subject is standing in the anatomical position (Figure 1-1): that is, standing erect with (1) the toes pointed forward, (2) the eyes directed to the horizon, (3) the arms by the sides, and (4) the palms of the hands facing forward.
ANATOMICAL CUTS AND PLANES
There are two basic ways to visualize deep structures of the human body. One is to dissect down to the area of interest; the other is to cut through the cadaver in defined planes (see Figure 1-1). Sections also can be obtained from a living patient with CT or MRI.
TERMS OF RELATIONSHIP
The following terms are presented in pairs because each term has an opposite (Figure 1-2). Again, the assumption is that our subject is in the anatomical position.
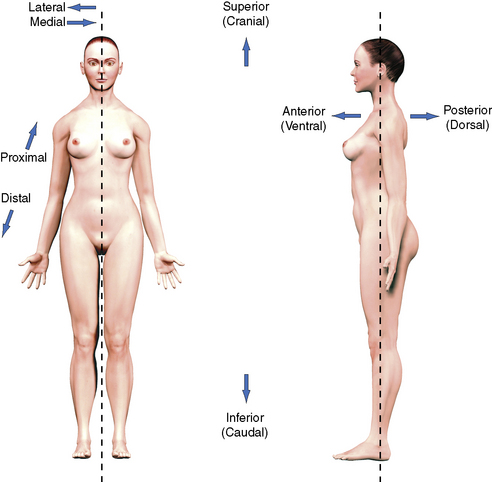
| Term | Definition |
|---|---|
| Anterior (ventral) | Toward the front of the body |
| Posterior (dorsal) | Toward the back of the body |
| Superior (cranial) | Toward the top of the head |
| Inferior (caudal) | Toward the soles of the feet |
| Medial | Toward the median plane |
| Lateral | Away from the median plane |
| Proximal (central) | Toward the trunk |
| Distal (peripheral) | Away from the trunk |
| Superficial | Toward the skin or body surface |
| Deep | Toward the interior of the body |
| Ipsilateral (homolateral) | On the same side |
| Contralateral | On the opposite side |
| Palmar surface of hand | Anterior surface of hand |
| Dorsal surface of hand | Posterior surface of hand |
| Plantar surface of foot | Inferior surface of foot |
| Dorsal surface of foot | Superior surface of foot |
3 Skeleton
CARTILAGE
Types
BONE
Functions
Classification
By Region
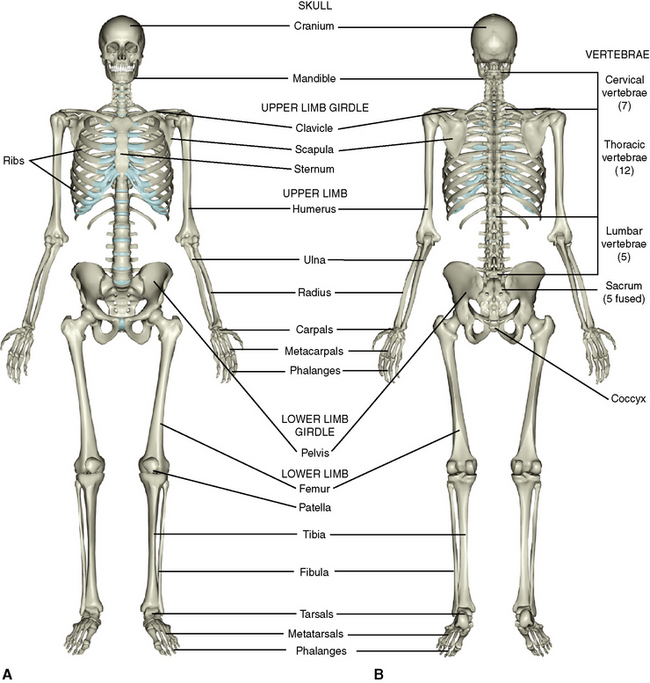
The adult skeleton is divided into an axial and an appendicular skeleton. The axial skeleton comprises the skull, the vertebral or spinal column, the ribs, and the sternum. The appendicular skeleton includes the bones of the upper and lower limbs. The individual bones and their numbers are illustrated in Figure 1-3.
By Shape
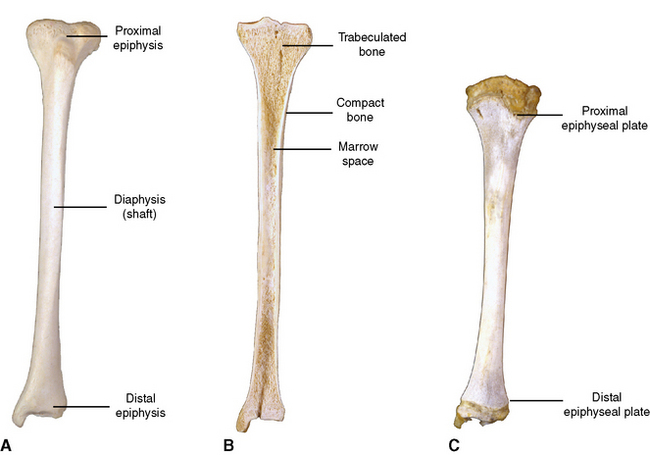
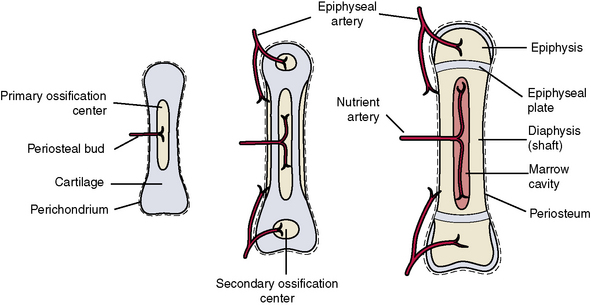
Long bones are hollow tubes, shafts, or diaphyses that are capped at both ends by knoblike epiphyses. A section through a long bone (Figure 1-4) reveals (1) an outer compact layer for rigidity, (2) an inner cancellous or spongy layer consisting of trabeculated bone for inner support, and (3) a marrow space containing blood cell–forming tissues in active red marrow or just plain fat in inactive yellow marrow.
The blood supply to long bones (Figure 1-5) is from the following three different sources: (1) nutrient arteries pierce the shaft and supply all layers to the marrow cavity within, (2) periosteal arteries supply periosteum and some adjacent compact bone, and (3) epiphyseal arteries supply the epiphyses and the adjacent joint structures.
Surface Features
Raised Markings or Elevations
Development
Endochondral Ossification
The remainder of the skeleton undergoes a slightly more complicated process of endochondral ossification (see Figure 1-5). Each of these bones is preformed in cartilage during early embryonic development. During the sixth to eighth weeks of embryonic development, cartilage within the center of the future bone shaft dies and is replaced by invading osteoblasts that form the primary center of ossification. The perichondrium surrounding the shaft becomes periosteum and it lays down an intramembranous collar of bone around the primary center. All the primary centers develop before birth. Invading vascular tissue hollows the shaft to form the medullary cavity that contains red bone marrow.
4 Joints
BY JOINT TISSUES
Fibrous Joints
There are three types of fibrous joints: (1) suture, (2) syndesmosis, and (3) gomphosis.
Sutures are found only between the bones of the skull (Figure 1-6). In the fetal skull the sutures are wide, and the bones present smooth opposing surfaces. This spacing between the flat bones of the skull allows a slight degree of movement between the skull bones during passage of the head through the birth canal (birth molding).
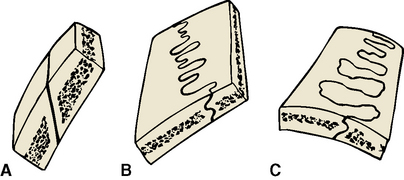
After birth, the sutures become quite rigid (synarthrodial) during infancy and early childhood, allowing no movement between skull bones. The developing sutures differentiate into one of three types (see Figure 1-6): (1) a squamous suture in which the bones simply overlap obliquely but are rendered immobile by intervening fibrous tissue; (2) a serrated suture, which develops sawtoothed interdigitating projections from the opposing bones; and (3) a denticulate suture, which features interlocking dove-tailed surfaces.
A syndesmosis, unlike other fibrous joints, is partially movable (amphiarthrodial) and is a joint in which the two bony components are farther apart, united by a fibrous interosseous membrane (Figure 1-7, A). Examples are the joint between the two bones of the forearm (radius and ulna) and the joint between the bones of the leg (fibula and tibia). Syndesmoses also are found between the laminae of the vertebrae.
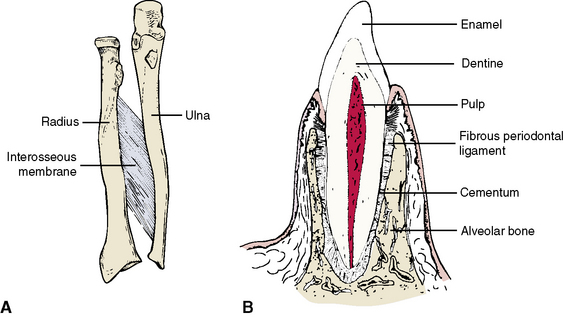
A gomphosis is a unique joint in the form of a peg-and-socket articulation between the roots of the teeth and the maxillary and mandibular aleveolar processes (Figure 1-7, B). Fibrous tissue organized as the periodontal ligament anchors the tooth securely in the socket. Mobility of this joint indicates a pathological state affecting the supporting structures of the tooth.
Primary Cartilaginous Joints (Synchondroses)
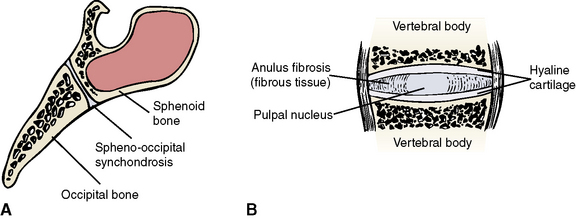
Primary cartilaginous joints develop between two bones of endochondral origin. They are characterized by a solid plate of hyaline cartilage between apposing surfaces (Figure 1-8, A). The cartilage plate functions in exactly the same manner as the epiphyseal plate between primary and secondary centers of long bones and provides an area of growth between bones. An example is the sphenooccipital synchondrosis in the young skull between the sphenoid bone and the occipital bone of the skull. This joint fuses after adolescence.
Secondary Cartilaginous Joints (Symphyses)
A secondary cartilaginous joint, or symphysis, is a partially movable (amphiarthrodial) joint in which the apposing bony surfaces are covered with cartilage but separated by intervening fibrous tissue or fibrocartilage (Figure 1-8, B). Symphyses are found in the midline of the body and include the joints between vertebral bodies (intervertebral discs), between right and left pubic bones (symphysis pubis), and in the newborn skull between the right and left halves of the mandible (symphysis menti). The symphysis menti starts to fuse during the first year of life to form a single bone, the mandible.
SYNOVIAL JOINTS
A synovial joint is freely movable (diarthrodial) and is typical of nearly all the joints of the upper and lower limbs. Synovial joints have a number of characteristic features (Figure 1-9, A).
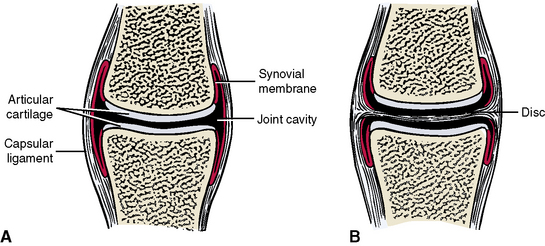
Articular cartilage coats the surfaces of the apposing bones. The cartilage may be hyaline (where bones of endochondral origin articulate) or may be fibrocartilage (where bones of intramembranous origin articulate). Typical of cartilage, this layer contains no blood vessels or nerves but must instead be nourished from the epiphyseal vessels of the bone and derives nourishment from the synovial lubricating fluid within the joint. A joint cavity exists between the articular surfaces of the apposing bones. The joint cavity is not large but contains enough space to allow a thin intervening film of synovial fluid. A capsular ligament surrounds the joint like a fibrous sleeve and attaches to the circumference of both bones to completely enclose the joint cavity. A synovial membrane consisting of loose areolar tissue contains a rich supply of capillaries. This membrane lines the inner aspect of the capsular ligament but does not line the articular surfaces of the cartilage. The synovial membrane secretes a lubricating synovial fluid, or synovium, into the joint cavity. Some joints contain discs interposed between the articular surfaces (Figure 1-9, B). A disc or meniscus is a fibrocartilaginous or sometimes condensed fibrous structure found within some joint cavities. These padlike structures divide the joint cavity into two compartments allowing for two types of movements, one for each subdivided joint compartment. The temporomandibular, or jaw, joint is an example of a synovial joint containing a disc.
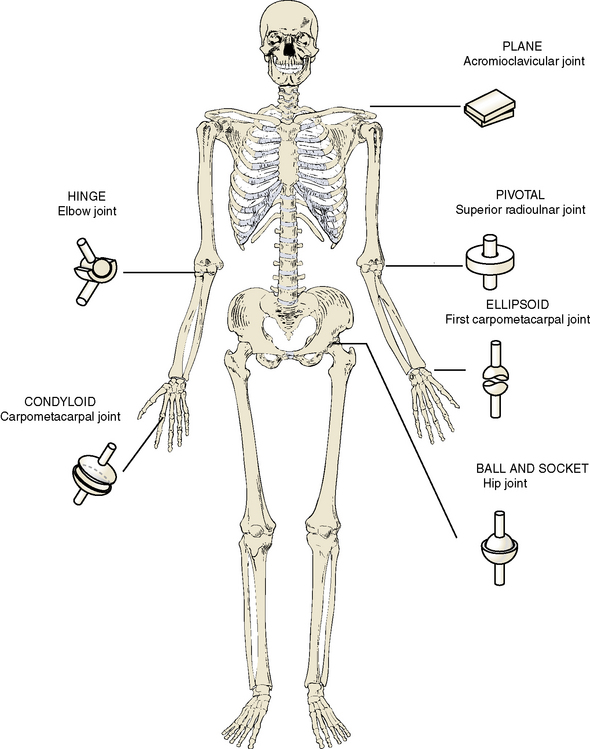
Classification
Freely movable synovial joints may be classified in different ways (Figure 1-10). One classification is based on the number of axes in which a joint can be moved (i.e., uniaxial, biaxial, or multiaxial). The shape or form of the opposing bony surfaces determines the degree of movement.
5 Muscular System
SKELETAL MUSCLE
Parts
Skeletal muscle consists of a fleshy portion and a fibrous or tendinous portion.
Fleshy Portion
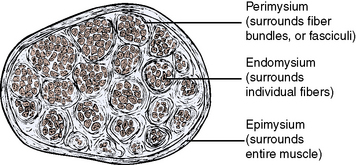
The cells derive their individual blood and nerve supply via sheets of fibrous connective tissue membranes through which the vessels and nerves run. Surrounding the individual muscle cells is a fibrous sheet of endomysium. Surrounding a bundle of several fibers (fasciculus) is a fibrous sheet of perimysium. Finally, surrounding several bundles, there is an overall fibrous coating of epimysium covering the entire muscle. Each of the three levels of surrounding membranes is interconnected, allowing vessels and nerves entering the outer layer eventually to reach individual fibers (Figure 1-11).
Groups of muscles in the limbs can be bound into compartments by intermuscular septa, which separate various groups of muscles. An example is the anterior compartment of the arm containing flexor muscles and the posterior compartment of the arm containing extensor muscles (see Chapter 9).
Stay updated, free dental videos. Join our Telegram channel
VIDEdental - Online dental courses


