Enamel, Dentin, and Pulp
• To discuss the changes in the inner enamel epithelial cells that allow them to become enamel-forming cells
• To discuss the interrelationship between enamel formation and dentin formation
• To describe the role of the dental papilla in the formation of the enamel organ and the shaping of the crown
• To describe the properties of enamel and the makeup of the enamel rod
• To understand the keyhole shape of the enamel rod and the direction of the hydroxyapatite crystals in different areas of the cross section of rod
• To define the following terms: striae of Retzius, hypoplastic enamel, hypocalcified enamel, enamel lamellae, enamel tuft, and enamel spindle
• To describe the properties and components of dentin
• To differentiate primary, secondary, and reparative dentin
• To define the following terms: interglobular dentin, dead tracts, sclerotic dentin
DENTAL PAPILLA
During the bud stage, the cells of the embryonic connective tissue deep to the bud resemble large multipointed cells called mesenchymal cells. As the enamel organ goes into the cap stage, the mesenchymal cells adjacent to the cap become more rounded and condensed and are then called dental papilla cells. The enamel organ is also enlarging at this time (Fig. 20-1). This condensation continues into the bell stage, during which further changes occur. It is at this point that the relationship between enamel and dentin formation becomes more obvious. Following is a list of events that occur during their formation:
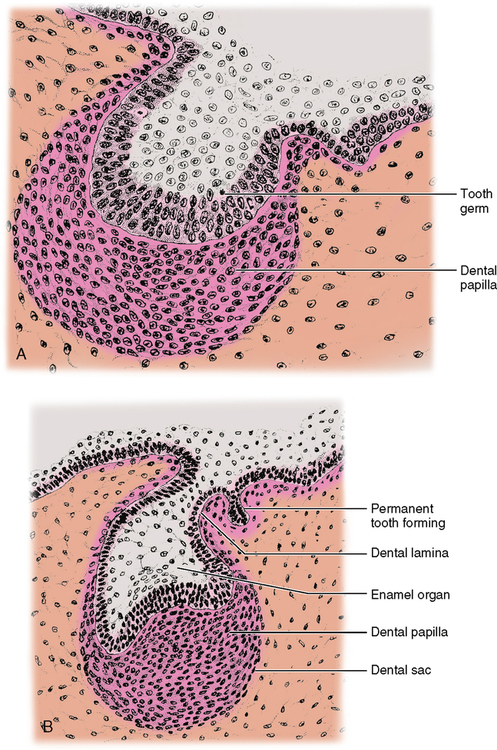
1. During the bell stage the inner enamel epithelial cells (IEE) become taller. They increase from 12 to 40 μm in length. These taller cells are then referred to as preameloblasts. As these cells enlarge, preparing to become secretory cells, their organelles are multiplying to prepare for this. We could expect to see more mitochondria and endoplasmic reticulum, and a more obvious Golgi apparatus.
2. The peripheral cells of the dental papilla adjacent to the preameloblasts become low columnar or cuboidal cells and are called odontoblasts (Fig. 20-2).
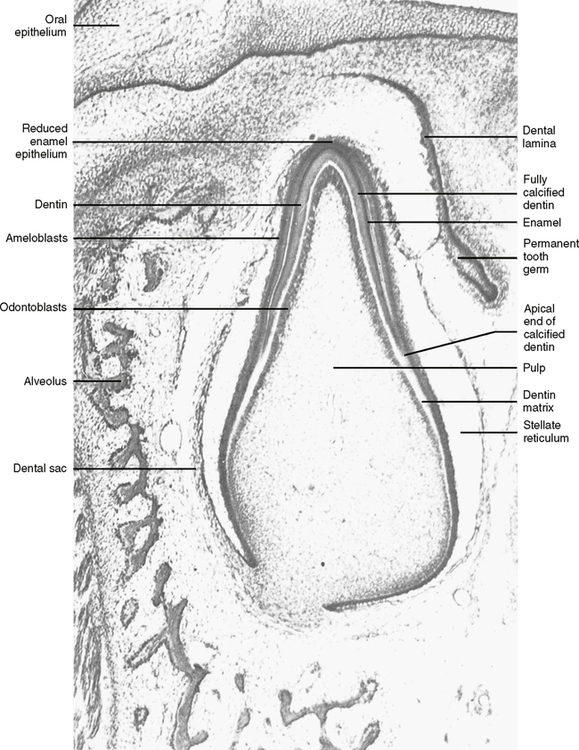
3. The odontoblasts move away from the preameloblasts toward the center of the dental papilla and secrete a matrix of mucopolysaccharide ground substance and collagen fibers.
4. The secretion of this dentin matrix causes the preameloblast to change its polarity (the nucleus moves from the center of the cell to the end nearest the stratum intermedium). This is attributable to the change in the route of nourishment of the preameloblasts. Until this time the nourishment had been going from the dental papilla directly to the adjacent cells. However, after the dentin matrix is laid down, it acts as a barrier between the dental papilla and the preameloblasts, and it becomes necessary for the preameloblasts to find a new route for nourishment. Vascular channels begin to penetrate the enamel organ through the outer enamel epithelium. Fluid nutrients pass to the stellate reticulum, the stratum intermedium, and then the preameloblasts. The nuclear shift of a preameloblast cell relates to the nucleus’s need to be closer to the nutrient supply. With the change in polarity the cell is called an ameloblast and is ready to begin the secretion of enamel matrix.
5. The ameloblast lays down a matrix of mucopolysaccharide and organic fiber next to the dentin matrix, and the future dentinoenamel junction (DEJ) is formed. As the ameloblast secretes the matrix, it moves away from the dentin toward the outer enamel epithelium (OEE) (Fig. 20-3).
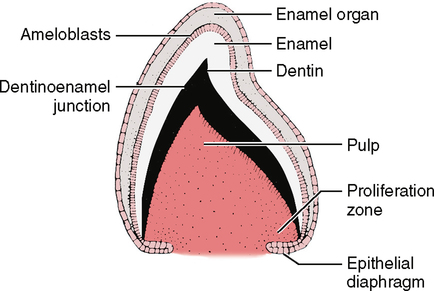
6. The dentin begins to lay down hydroxyapatite crystals and calcify (crystals begin growing).
7. The enamel begins to lay down hydroxyapatite crystals and calcify (crystals begin growing).
ENAMEL COMPOSITION
The enamel structure comprises two parts: the rod sheath and the enamel rod. The rod sheath outlines the rod and contains most of the fibrous organic substance (Fig. 20-4). However, the rod, which is made up of hydroxyapatite crystals, is the primary unit of enamel’s structure. The rod is a column of enamel that runs all the way from the dentinoenamel junction to the surface of the tooth. It is somewhat perpendicular to the dentinoenamel junction and to the surface of the crown (Fig. 20-5).
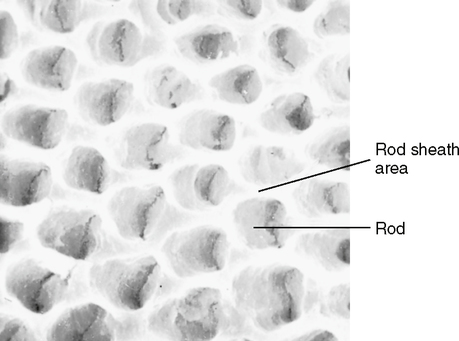
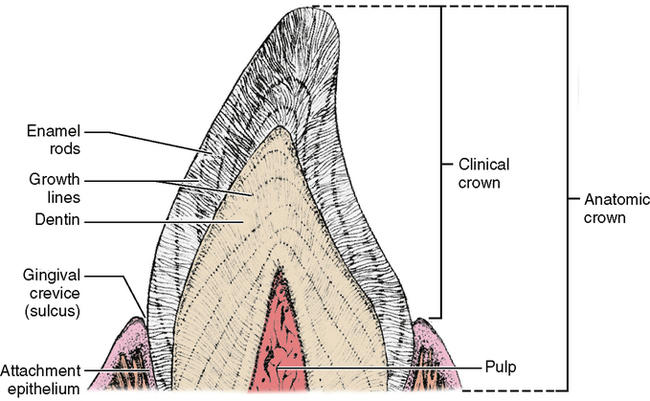
A “typical” enamel rod with the keyhole shape (Fig. 20-6, A) has a wide upper end and a narrowed bottom end. The reason for the different appearances of the upper and lower parts of the keyhole is that the crystals in those parts are oriented in different directions. When the ameloblasts produce the matrix and then deposit the crystals into the matrix, the secreting ends of the ameloblasts bulge outward in the direction of the DEJ. This asymmetrical bulge is called the Tomes’ process of an ameloblast. Each ameloblast does not lie parallel to the long axis of the enamel rod but is canted at an angle. As the crystals are secreted, they emerge from the pointed part of the ameloblast in an arrangement perpendicular to the cell membrane. The result is that the crystals in the upper part of the keyhole are arranged in a pattern longitudinal to the long axis of the rod, while the crystals in the lower end are angled almost 60 degrees off of the long axis.

Stay updated, free dental videos. Join our Telegram channel
VIDEdental - Online dental courses


