Temporomandibular Joint
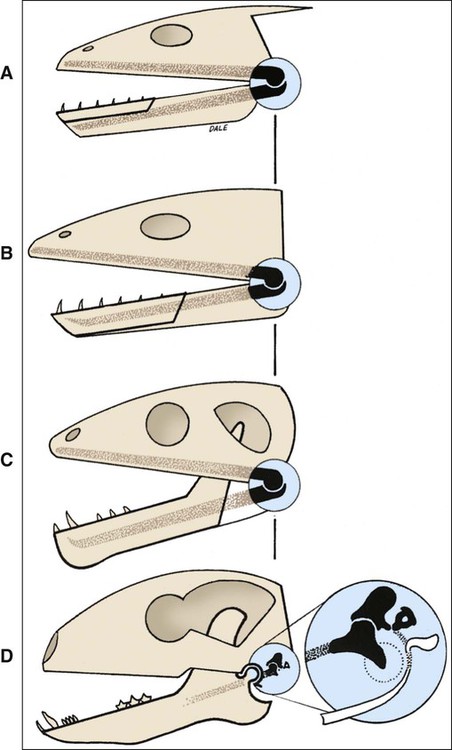
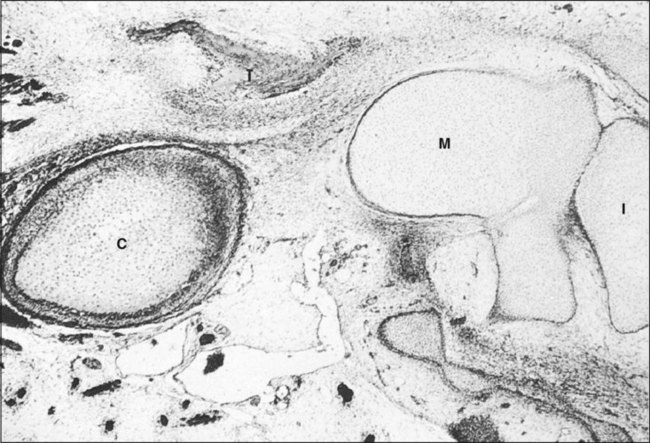
The bones involved of the articulation of the lower jaw with the cranium and upper facial skeleton are the mandible and the temporal bone, and the joint therefore is designated the temporomandibular joint (TMJ). The joint is unique to mammals. In other vertebrates the lower jaw is compound, consisting of several bones including the dentary bone (bearing teeth) and the articular bone (formed from the posterior part of Meckel’s cartilage), and articulates with the quadrate bone of the skull (Figure 13-1). As mammals evolved, the compound lower jaw was reduced to a single bone (the mandible) bearing teeth that articulate with the newly developed articulating surface on the temporal bone. Thus in phylogenetic terms the TMJ is a secondary joint. The primary vertebrate jaw joint is still present in human anatomy (as the incudomalleolar articulation), with the bones involved (incus and malleus) now positioned in the middle ear (Figure 13-2).
Type of Joint
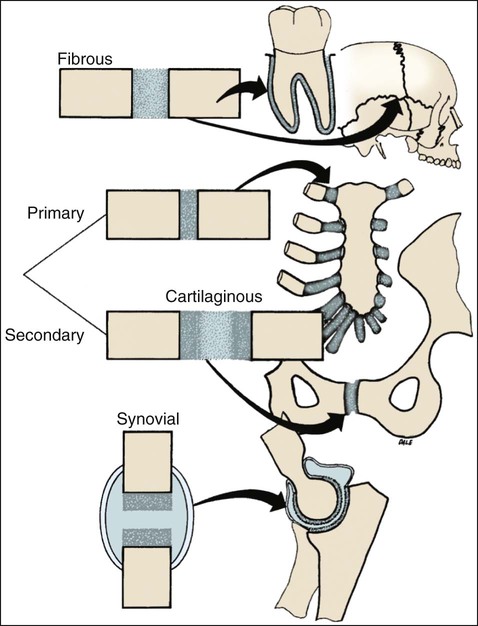
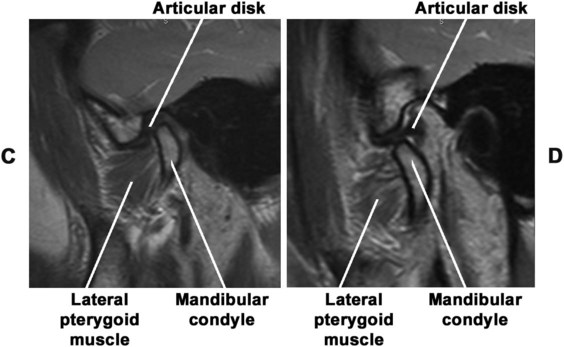
The temporomandibular articulation is a synovial joint. The anatomy of the TMJ varies considerably among mammals, depending on masticatory requirements, so that a single, all-embracing descriptive classification is not possible. In carnivores, for example, movement is restricted to a simple hinge motion by the presence of well-developed anterior and posterior bony flanges that clasp the mandibular condyle. The badger provides an extreme example of this—the flanges clasp and envelop the condyle to such an extent that it is not possible to dislocate the mandible from the skull. In human beings a different situation exists; the masticatory process demands that the mandible be capable not only of opening and closing movements but also of protrusive, retrusive, and lateral movements and combinations thereof. To achieve them, the condyle undertakes translatory and rotary movements; therefore, the human TMJ is described as a synovial sliding–ginglymoid joint (Figure 13-4).
Development of the Joint
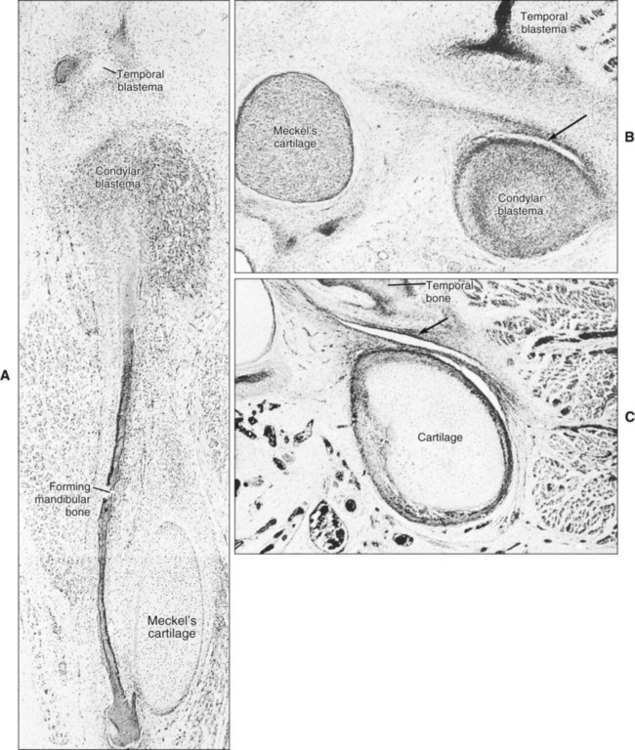
At 3 months of gestation, the secondary jaw joint, the TMJ, begins to form. The first evidence of TMJ development is the appearance of two distinct regions of mesenchymal condensation, the temporal and condylar blastemata. The temporal blastema appears before the condylar, and initially both are positioned some distance from each other. The condylar blastema grows rapidly in a dorsolateral direction to close the gap. Ossification begins first in the temporal blastema (Figure 13-5, A). While the condylar blastema is still condensed mesenchyme, a cleft appears immediately above it that becomes the inferior joint cavity (Figure 13-5, B). The condylar blastema differentiates into cartilage (condylar cartilage), and then a second cleft appears in relation to the temporal ossification that becomes the upper joint cavity (Figure 13-5, C). With the appearance of this cleft, the primitive articular disk is formed.
Bones of the Joint
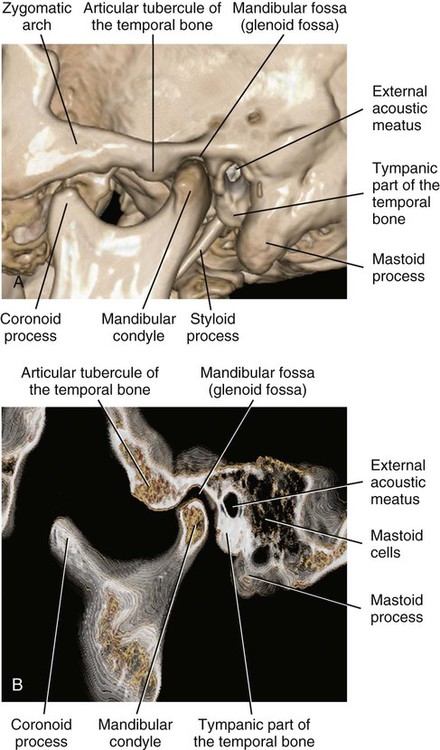
The bones of the temporomandibular articulation are the glenoid fossa (on the undersurface of the squamous part of the temporal bone) and the condyle (supported by the condylar process of the mandible). The glenoid fossa is limited posteriorly by the squamotympanic and petrotympanic fissures. The glenoid fossa is limited medially by the spine of the sphenoid and laterally by the root of the zygomatic process of the temporal bone. Anteriorly, the glenoid fossa is bounded by a ridge of bone described as the articular eminence, which also is involved in the articulation (Figure 13-6). The middle part is a thin plate of bone, the upper surface of which forms the middle cranial fossa (housing the temporal lobe of the brain). The condyle is the articulating surface of the mandible. Viewed sagittally, the glenoid fossa is 15 to 20 mm long (from medial to lateral extreme) and 8 to 12 mm thick. The articular surface of the condyle is strongly convex in the anteroposterior direction and slightly convex mediolaterally. The medial and lateral ends are termed poles. The medial pole extends farther beyond the condylar neck than the lateral pole does and is positioned more posteriorly so that the long axis of the condyle deviates posteriorly and meets a similar axis drawn from the opposite condyle at the anterior border of the foramen magnum. Variations in the shape of the condyle are frequent, and often the condylar surface is divided by a sagittal crest into medial and lateral slopes.
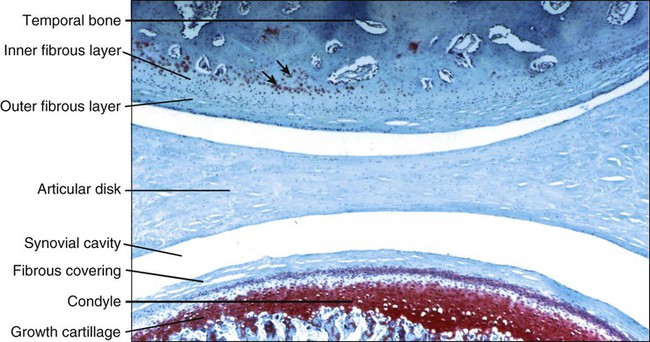
Unlike most synovial joints, the articular surfaces of which are covered with hyaline cartilage, the temporomandibular articulation is covered by a layer of fibrous tissue (Figure 13-7). This histologic distinction has been used to argue that the TMJ is not a weight-bearing joint, but the reality for this distinction can be found in the developmental history of the joint. The only other synovial joints with articular surfaces covered by fibrous tissue are the acromioclavicular and sternoclavicular, linking the clavicle to the appendicular skeleton. The mandible and the clavicle are bones formed directly from an intramembranous ossification center and are not preformed in cartilage, cartilage that persists in the long bones to cover articular surfaces following the appearance of ossification centers.
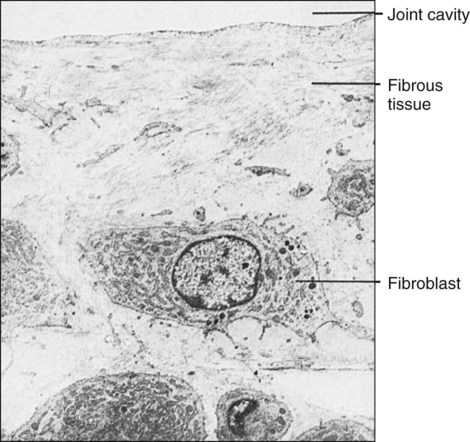
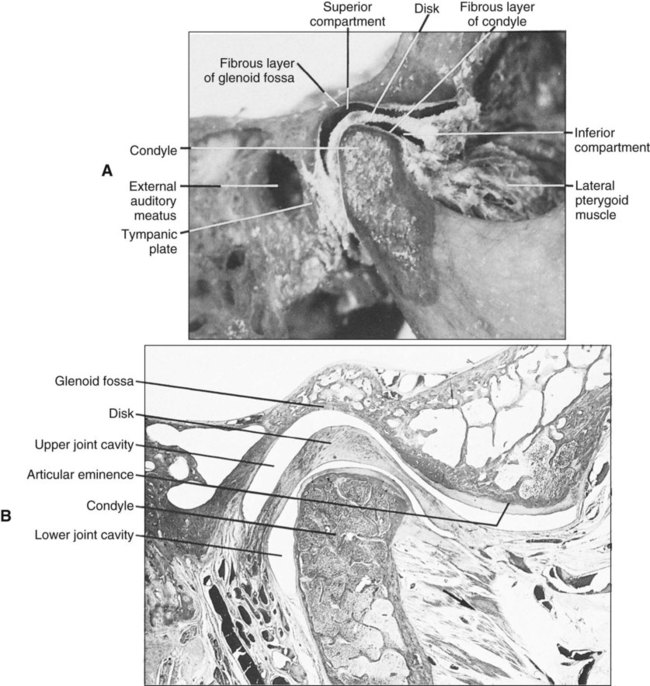
The glenoid fossa always is covered by a thin fibrous layer that directly overlies the bone, much as periosteum does (Figure 13-8), but this layer becomes appreciably thicker where it covers the slope of the articular eminence (Figure 13-9).
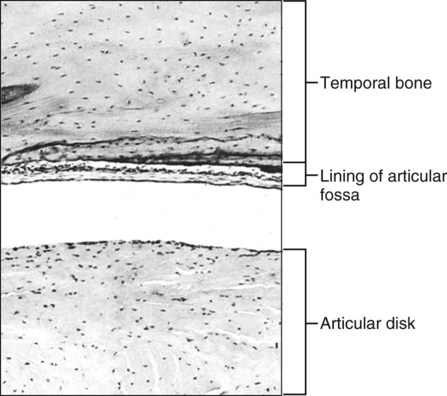
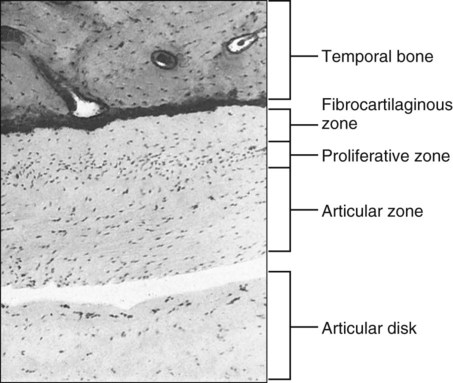
Cartilage Associated with the Joint
Earlier accounts of TMJ histology indicated that the surface coverings of the joint consist of fibrocartilage rather than fibrous tissue. Although with age the fibrous covering layer might contain some cartilage cells, no evidence indicates that this is normal. However, firm evidence indicates that fibrocartilage is associated with the articulation deep to the fibrous layer, in the condyle and on the articular eminence (Figure 13-10). The occurrence of such cartilage has a developmental explanation: a secondary growth cartilage associated with the developing TMJ forms within the blastema the condylar cartilage and is in some ways akin to the epiphyseal cartilage of a developing long bone. The condylar cartilage consists essentially of a proliferative layer of replicating cells that function as progenitor cells for the growth cartilage (Figure 13-11). These cells become chondroblasts and elaborate proteoglycans and type II collagen to form the extracellular matrix of cartilage, in which they become entrapped as chondrocytes. At the same time, an increase in the size of the chondrocytes occurs (hypertrophy). Following the production of this cartilage, endochondral ossification occurs and involves mineralization of the cartilage, vascular invasion, loss of chondrocytes, and differentiation of osteoblasts to produce bone on the mineralized cartilaginous framework (Figure 13-12; see also Figure 13-11). The only difference in this process between condylar and epiphyseal cartilages in long bones is the absence of ordered columns of cartilaginous cells (which characterize the epiphyseal growth cartilage and results from chondroblast cell division). The absence of well-defined, elongated columns of chondroblast daughter cells in condylar cartilage has key significance. A typical long bone epiphyseal plate characterized by well-defined columns is committed to an essentially unidirectional mode of growth; that is, the proliferation of cells by mitotic division is such that the whole bone necessarily elongates in a manner determined by the columns of dividing cells. The mandibular condyle, by contrast, has a multidirectional growth capacity, and its cartilage can proliferate in any combination of superior and posterior directions as needed to provide for the best anatomic placement of the mandibular arch.
Stay updated, free dental videos. Join our Telegram channel
VIDEdental - Online dental courses


