Providing motion is the central function of the craniomandibular or temporomandibular articulation, where mandible and cranium are connected as the temporomandibular joint (TMJ). As part of the stomatognathic system, the TMJ allows simple and complex movements such as free movement during phonation, gesticulation, swallowing, and breathing, as well as different types of load-bearing movements during incision and chewing. Although all other bony segments of the head are fused via synostoses, only the three ossicles within the middle ear also show articulations. Therefore, the description of the phylogenetic development of the TMJ is helpful for an understanding of its structure and function.
DEVELOPMENTAL PERSPECTIVE OF THE CRANIOMANDIBULAR ARTICULATION
In mammals, the articulating bony parts of the TMJ are derived from membrane bone. These are formed directly from intramembranous centers of ossification. These developing bones, the squamous portion of the temporal bone, and the condyles of the mandible become completely surrounded by periosteum, including areas that eventually form the articular surfaces of the TMJ and build the “secondary” TMJ there. In contradistinction to that is the development of the “primary” TMJ of nonmammalians, deriving from cartilage. In bony fish, reptiles, and birds, the proximal parts of the mandible constitute the TMJ, which is actually the primary TMJ. In mammals, the primary or primitive TMJ is subjected to complex changes, also ultimately forming the joints for three ossicles within the middle ear. The secondary TMJ in humans starts to develop by the 10th week of gestation from mesenchymal condensations of two separate blastemas for the mandibular condyle and the temporal bone. The condylar and temporal mesenchymal cells differentiate into osteoblasts, which lay down membrane bone. The periosteum that lines these articular surfaces gradually is transformed during its early development into the dense fibrous articular tissues of the TMJ. In the center of the condyle, cartilage develops and becomes the growth center, with subchondral bone formation by an enchondral mechanism. Superior to the condylar blastema, a band of mesenchymal cells develops that differentiates into the disc, which is considered to derive as an extension of lateral pterygoid muscle. The configuration and shape of the TMJ are also related to the development of dentition. During infancy, the articular fossa is a flat furrow with almost no eminence. Eruption of the primary dentition increases the groove, forming a fossa. The final adult formation of the TMJ is associated with eruption of the permanent dentition. Articular forces acting through the TMJ play an important role in this gradual transformation. Depending on the preservation of the dentition, the TMJ may maintain a stable form throughout life. Edentulism of the jaws again causes secondary changes within the TMJ with flattening of the articular fossa and eminence. The configuration and shape of the TMJ depend on skeletal and dental relationships of the maxilla and mandible. For example, a deep bite is correlated with a steep eminence.
Most synovial joints have hyaline cartilage that lines their articulating surfaces because bones of typical synovial joints are endochondrally formed bones that are initially pre-formed in hyaline cartilage. In contrast, articulating surfaces of the TMJ are lined by dense, avascular, fibrous connective tissue. Most of this cartilage eventually is calcified and then is replaced by bone, although the cartilage that lines the articular surfaces persists in a modified form. Thus, the lack of hyaline cartilage on the articular surfaces of the TMJ reflects its unique ontogenetic and phylogenetic development; although it is not a hyaline cartridge–covered joint, it is capable of bearing loads. Although a secondary cartilage is present in the condyle of the growing mandible, this cartilage does not form part of the articular surface because the periosteum-derived articular tissues cover it.
SURGICAL ANATOMY AND HISTOLOGY
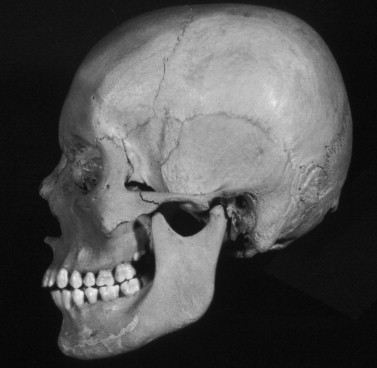
The mandible and cranium make up the craniomandibular articulation, which frequently is referred to as the temporomandibular articulation . This articulation of the mandible consists of two synovial joints: left- and right-sided TMJs. The bony components include the mandibular condyles below and the squamous temporal bones above ( Figure 42-1 ). The articular space of each TMJ is divided into upper and lower compartments because of the interposition of the fibrous articular disc between the temporal bone and the mandible. Gliding or translatory movements occur primarily in the upper compartment, while the lower compartment functions primarily as a hinge or rotary joint.
GLENOID FOSSA—ARTICULAR EMINENCE
The glenoid , or mandibular fossa , is the concavity within the temporal bone that houses the mandibular condyle. Its anterior wall is built by the articular eminence of the squamous temporal bone and its posterior wall by the tympanic plate of the temporal bone, which also forms the anterior wall of the external acoustic meatus. The bony roof of the glenoid fossa is thin and often appears translucent when transilluminated ( Figure 42-2 ).
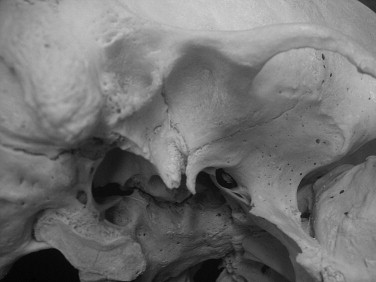
It is important to note that the term articular fossa refers to the particular portion of the glenoid fossa that is covered by articular tissues. It is built entirely by the squamous portion of the temporal bone (see Figure 42-1 ). The posterior part of the articular fossa is elevated to a ridge called the posterior articular lip . In most individuals, the posterior articular lip is higher and thicker at its lateral end and thus is visible from the side as a cone-shaped process between the articular fossa and the tympanic plate. This structure is the postglenoid process. The lateral border of the articular fossa sometimes is marked by a narrow, low ridge or crest.
In the posterior and lateral parts of the glenoid fossa , a fissure separates the articular and nonarticular portions of the glenoid fossa. This fissure, called the tympanosquamosal fissure , also separates the tympanic portion from the squamous portion of the temporal bone. Medial to this fissure, a bony plate of the petrous portion of the temporal bone, the tegmen tympani, protrudes between the tympanic and squamous portions. Therefore, instead of a tympanosquamosal fissure along the medial aspect of the glenoid fossa, an anterior petrosquamosal fissure and a posterior petrotympanic fissure occur. The petrotympanic fissure is slightly widened laterally to permit passage of the chorda tympani nerve and the anterior tympanic blood vessels. These neurovascular structures are located within the glenoid fossa, but not within the articular fossa (see Figure 42-2 ).
The articular eminence is the transverse bar of dense bone that forms the posterior root of the zygomatic arch and the anterior wall of the articular fossa. It has a large articular surface. It is strongly convex on a side view and moderately concave when viewed from the front or back. The degree of this convexity and concavity is highly variable. Fine bony ridges often outline the medial and lateral borders of the articular eminence. Although a thin layer of fibrous tissue covers the roof of the articular fossa, the fibrous tissue that covers the articular eminence is thick and quite firm. Moreover, different from the roof, the articular eminence is made up of a fairly thick layer of dense bone. Lateral to the articular eminence, a small bony projection, the articular tubercle , is located. The articular tubercle is not an articular surface. Instead, it serves as the attachment area for portions of the temporomandibular ligament (TML).
ARTICULAR DISC—DISC LIGAMENTS
The articular disc is a firm, oval, fibrous plate that is positioned between the mandibular condyle and the articular fossa and eminence ( Figure 42-3 ). Its central part, the intermediate zone, is considerably thinner than its periphery—the anterior and posterior bands. Anteriorly, the disc continues as the anterior attachment and is fused to the capsule of the TMJ. Posteriorly, the disc continues as the posterior attachment or bilaminar zone—a thick double layer of vascularized connective tissue. The bilaminar zone splits into two parts: (1) an upper fibroelastic layer that attaches to the postglenoid process, posterior articular lip, and tympanosquamosal fissure; and (2) a lower fibrous layer that attaches to the posterior portion of the condylar neck immediately below the articular tissues. Posteriorly, these two layers are separated by the intermediate layer, which contains loose connective tissue that attaches to the posterior wall of the joint capsule. The posterior attachment has a profuse supply of nerves and blood vessels.
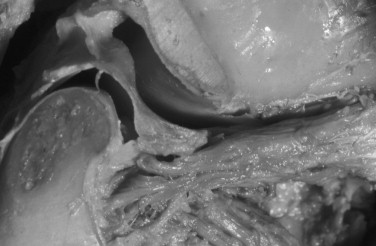
Unlike its anterior and posterior attachments, the disc is not attached to the capsule laterally or medially. Instead, it is tightly bound directly to the medial and lateral poles of the mandibular condyle. The position of the disc relative to the condyle also is influenced by the pull of the superior head of the lateral pterygoid muscle because a small portion of this muscle often attaches to the disc ( Figure 42-4 ). Because the superior head of the lateral pterygoid also attaches to the mandibular condyle, and the disc is tightly bound to the medial and lateral poles of the condyle, this muscle arguably may have no special influence on movements of the articular disc relative to the condyle.

Ontogenetically, the articular disc derives from a mesenchymal block of tissue that also gives rise to the capsule of the TMJ and the lateral pterygoid muscle. This tissue mass is located between the developing squamous temporal bone and mandibular condyle. In adults, the uppermost part (or superior head) of the lateral pterygoid muscle often retains its original connection to the capsule and articular disc of the TMJ.
Blood vessels and nerves are absent in the intermediate zone, that is, the firm central region of the articular disc, as well as in the avascular fibrous layers that cover the mandibular and temporal articular surfaces of the joint. A lack of these neurovascular structures is compatible with the hypothesis that a considerable reaction force occurs along this portion of the joint. It has been suggested that one main function of the articular disc is to reduce stress concentrations between the articular surfaces of the mandibular condyle and the squamous temporal bone, that is, the compliant nature of the disc helps to distribute reaction force more evenly along these joint surfaces.
CONDYLE
The articular surface of the mandible is the upper and anterior surface of the condyle ( Figure 42-5 ). The adult human condyle is about 15 to 20 mm from side to side and 8 to 10 mm from front to back. Its long axis lies at right angles to the plane of the mandibular ramus. Because of the flare of the ramus, however, the long axes of the left and right condyles cross approximately at the anterior margin of the foramen magnum, forming an obtuse angle varying from 145 to 170 degrees.
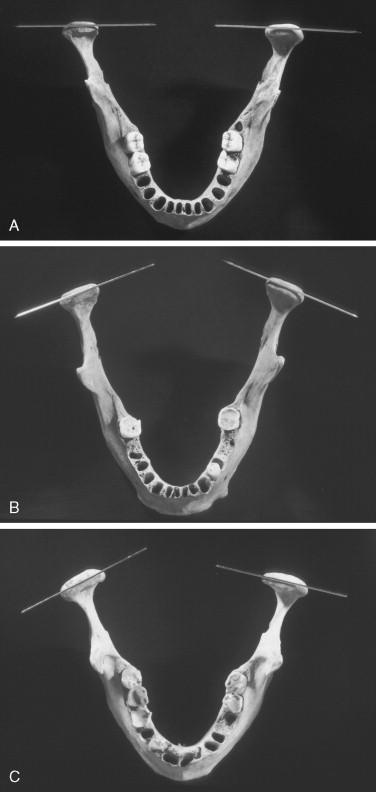
The articular surface of the condyle is strongly convex when viewed from the side and less so when viewed from the front. The articular surface faces upward and forward, so that in side view, the condylar neck is bent forward. As seen from the front, the articular convexity often resembles a tent-like configuration that is divided into medial and lateral slopes by a variably prominent crest. The lateral pole of the condyle extends slightly beyond the outer surface of the ramus and is roughened for the attachment of the articular disc and the TML. The medial pole of the condyle protrudes considerably beyond the inner surface of the ramus and also is slightly roughened for the attachment of the articular disc. Variations in the shape of the condyle are frequent. Irregularities of the bony articular surfaces are obscured and smoothed by the thick covering of fibrous tissue that is derived from and directly continuous with the periosteum of the mandible.
SYNOVIAL MEMBRANE
The synovial membrane, a highly vascularized layer of connective tissue, lines all structures of the articulation that do not experience compressive forces. The largest area of synovial lining covers the upper and lower surfaces of the posterior attachment, including the loose connective tissue that binds the posterior border of the disc to the capsule. Synovial tissue also lines the inner aspect of the fibrous capsule. When the condyle is positioned in the glenoid fossa, the synovial membrane forms rather heavy folds posteriorly (see Figure 42-4 ). When the condyle is protruded toward the summit of the articular eminence, the folds disappear as the synovial tissues are stretched.
CAPSULE—EXTRACAPSULAR LIGAMENTS
The articular capsule is quite thin anteromedially, medially, and posteriorly, but it is thick anterolaterally and laterally, where it attaches to the articular tubercle. This reinforced lateral portion of the capsule is the temporomandibular ligament (TML). The capsule attaches to the squamous portion of the temporal bone along the outer limits of the articular surface of the articular eminence, fossa, and preglenoid plane. Posteriorly, the capsule arises from the postglenoid process, the posterior articular lip, and the tympanosquamosal fissure ( Figure 42-6 ).
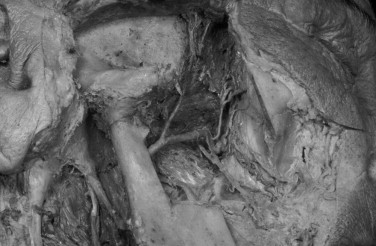
The anatomy of the capsule and that of the TML show considerable variability. The TML can be split into two layers: a wide, fan-shaped superficial portion, and a narrow deep portion. The broad origin of the superficial portion along the articular tubercle and its narrower insertion along the condylar neck account for its somewhat fan-shaped morphology. Its anterior fibers run from the articular tubercle obliquely down and back; the posterior fibers have a more vertical orientation. Fibers of the deep portion are oriented horizontally (anteroposteriorly), attaching along the lateral pole of the mandibular condyle and extending to a crest situated along the articular tubercle. This deep horizontal band sometimes is named the lateral polar ligament .
The fibrous capsule of the TMJ and the TML allow and limit jaw movements. The vertical fibers control distraction movements of the condyle from the articular eminence and fossa, the horizontal fibers (polar ligaments) prevent excessive retrusive movements of the condyle, and the posterior portion of the capsule limits protrusive movements. The anterior part of the capsule and the anterolateral part of the TML may limit condylar rotation during jaw opening, although most of this limitation is imposed by the stretched jaw-closing muscles.
ACCESSORY LIGAMENTS
The sphenomandibular and the stylomandibular ligaments are considered accessory ligaments of the temporomandibular articulation ( Figure 42-7 ).
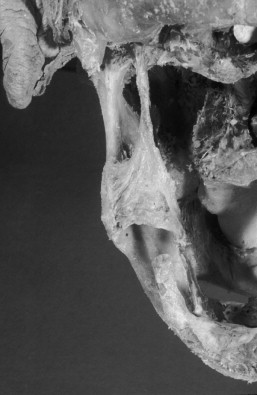
Sphenomandibular Ligament
The sphenomandibular ligament arises from the spine of the sphenoid bone and is directed downward and outward, inserting on the mandible at the mandibular lingula, which is located along the upper border of the mandibular foramen. It is derived from Meckel’s cartilage. In most individuals, the sphenomandibular ligament is a thin layer of connective tissue with indistinct anterior and posterior borders that protect the blood vessels and nerves passing through the mandibular foramen from additional tensile stress during jaw opening and closing. It normally has no influence on mandibular movement.
Stylomandibular Ligament
The stylomandibular ligament is a reinforced sheet of cervical fascia that extends from the styloid process and the stylohyoid ligament to the region of the mandibular angle. Many of its fibers are attached to the back edge of the lower part of the mandibular ramus; others continue onto the deep fascia along the medial surface of the medial pterygoid muscle. The upper border of the stylomandibular ligament normally is a thickened cord-like structure.
This ligament is relatively loose when the jaws are closed and when they are wide open; it is tensed only when the mandible is maximally protruded. Thus, apparently this ligament can limit excessive protrusive movements.
MUSCLES
Various muscle groups of the face, tongue, palate, and hyoid are involved in TMJ motion. In addition to the four powerful masticatory muscles (masseter, temporalis, medial pterygoid, and lateral pterygoid), the morphology and function of other important muscles that play a role in mandibular movements are described here.
Masseter Muscle
The masseter muscle reaches as a rectangular plate from the zygomatic arch to the lateral surface of the mandibular ramus ( Figure 42-8 ). It is divided into a superficial and a smaller deep portion. The superficial masseter arises from the lower border of the zygomatic arch as strong tendinous fibers. The most anterior fibers may arise from the outer corner of the zygomatic process of the maxilla. Posteriorly, the origin of the superficial portion ends along the zygomaticotemporal suture. The superficial masseter is covered on its outer surface by a strong tendinous layer that extends down from the zygomatic arch over the upper third or half of the muscle.

In a frontal view, these fibers are directed downward and medially. In a side view, the muscle fibers of the superficial masseter are directed downward and backward to insert along the angle of the mandible. The mandibular attachment of the superficial masseter extends along the lower one third of the posterior border of the ramus and along the lower border of the mandible anterior to the third molar; it covers, more or less, the lower half of the lateral surface of the ramus. The field of insertion has ridges into which the tendons insert, as well as grooves between the ridges into which the fleshy fibers insert.
Posteriorly, the fibers of the superficial masseter wrap around the posterior and inferior aspects of the angle of the mandible, joining fibers of the medial pterygoid muscle in a tendinous raphe. This muscular arrangement is called the pterygomasseteric sling . The deep and superficial portions of the masseter fuse anteriorly, but posteriorly, the two can be separated. Fibers of the deep masseter arise from the entire length of the zygomatic arch up to the anterior slope of the articular eminence. Some of its fibers also may arise from the lateral wall of the TMJ capsule.
The deep masseter inserts above the superficial masseter along the mandibular ramus as a triangular insertion field. The base of this triangle faces posteriorly, and the apex faces anteriorly. In the side view, the fibers of the deep masseter, which have a near vertical alignment, pass downward at an angle of about 30 to 40 degrees to the fibers of the more obliquely aligned superficial masseter.
The masseter muscle is a powerful elevator of the mandible. A lateral view reveals that the deep masseter exerts primarily a vertical force on the mandible. In contrast, the superficial masseter exerts a vertical and slightly anteriorly directed force on the mandible that is approximately perpendicular to the occlusal plane of the molars. The entire masseter also exerts a lateral component of force on the mandible.
Temporalis Muscle
The temporalis muscle has a fan-shaped origin along the lateral surface of the skull and the dense fascia overlying this muscle ( Figure 42-9 ). The bony attachment field, the temporal fossa, is encircled above by the inferior temporal line. This attachment field includes a narrow strip of the parietal bone, the greater part of the temporal squama, the temporal surface of the frontal bone, and the temporal surface of the greater wing of the sphenoid bone. Muscle fibers and tendons also arise from the postorbital septum, which is the bony partition that separates the temporal fossa from the orbit. Both the zygomatic and frontal bones and the greater wing of the sphenoid contribute to the formation of the postorbital septum. The bony field of origin of the temporalis muscle reaches downward to include the infratemporal crest of the sphenoid.
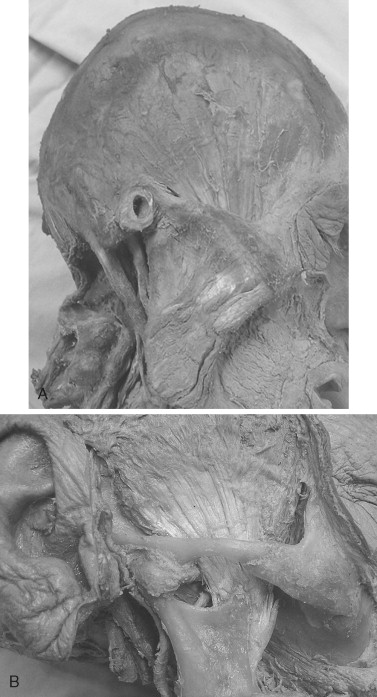
Many of the temporalis muscle fibers originate from the medial surface of the temporalis fascia. The temporalis fascia attaches to the superior temporal line and the upper border of the zygomatic arch. Passing downward from the superior temporal line, the temporalis fascia thickens considerably and then splits into two layers; the superficial layer continues into the periosteum of the zygomatic arch along its lateral surface, and the deep layer extends into the periosteum of the zygomatic arch along its medial surface.
The bundles of the temporalis muscle converge toward the opening located between the zygomatic arch and the lateral surface of the skull (temporal foramen). The tip of the coronoid process projects into this opening. The anterior fibers of the temporalis muscle, which form the major bulk of the muscle, are largely vertical; the fibers in the middle part of the muscle are increasingly oblique. The most posterior fibers run forward almost horizontally, bend around the posterior root of the zygomatic arch in front of the articular eminence, and pass downward vertically to the mandible.
The middle and posterior portions of the temporalis muscle, respectively, are attached along the apex of the coronoid process and along its posterior slope to the deepest point of the mandibular notch. The more superficial fibers of the anterior temporalis muscle insert along the apex of the coronoid process, the anterior surface of the coronoid process, and the mandibular ramus. The deeper fibers of the anterior temporalis attach along the medial anterior surface of the mandibular ramus.
Similar to the masseter muscle, the temporalis muscle mainly elevates the mandible. Its fan-shaped morphology indicates that its direction of pull varies considerably, depending on which portions are mechanically active. Superficially, it appears that its most posterior fibers retract the mandible because of their horizontal orientation along the side of the skull; however, as previously noted, when the condyle is situated in the mandibular fossa, the fibers of the posterior temporalis are bent around the posterior root of the zygomatic arch at a sharp angle and thus are oriented vertically. Therefore, this portion of the temporalis muscle exerts primarily an upward force on the mandible during normal closure. When the condyle is translated anteriorly into a more protruded position, these posterior fibers likely cause the mandible to retrude because, in this instance, the posterior temporalis is aligned more horizontally. Because its most posterior fibers pass very close to the condyle, the posterior temporalis probably also functions as a stabilizer of the TMJ.
The middle and obliquely aligned portion of the temporalis muscle is capable of exerting a vertical and retracting force on the mandible. Most of the anterior portion is capable of a vertical pull on the mandible. That portion of the anterior temporalis that originates from the postorbital septum, however, likely pulls the mandible upward and forward. Finally, the deep fibers of the anterior temporalis that originate along and just above the infratemporal crest pull the mandible upward and somewhat medially. Thus, the morphology of the entire temporalis muscle indicates that its fibers are capable of considerable variability in their direction of pull.
Medial Pterygoid Muscle
The medial pterygoid muscle is situated on the medial side of the mandibular ramus as a powerful rectangular muscle ( Figure 42-10 ). Its main origin is the pterygoid fossa, a depression that is located between the back edges of the medial and lateral pterygoid plates of the sphenoid bone. The deepest fibers arise from strong tendons; others arise directly from the medial surface of the lateral pterygoid plate. A flat tendon covers the medial surface of the muscle at its origin, and it is as wide as the tensor veli palatini, with which it is in contact.
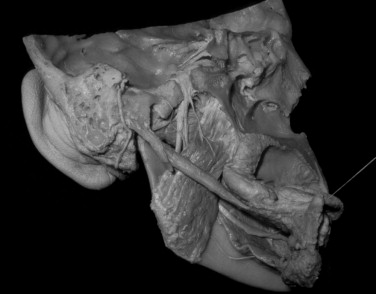
The most anterior fibers of the medial pterygoid arise from the outer and inferior surface of the pyramidal process of the palatine bone and from adjacent parts of the maxillary tuberosity. These fibers, which are referred to as the superficial head of the medial pterygoid, are positioned lateral to the lateral pterygoid muscle. The remaining and largest portion of this muscle, the deep head, is positioned medial or deep to the lateral pterygoid muscle.
The fibers of the medial pterygoid muscle run downward, backward, and laterally and are inserted along the medial surface of the angle of the mandible. The field of insertion is approximately triangular and is located between the mandibular angle and the mylohyoid groove. As noted earlier, the fibers of the medial pterygoid muscle often meet fibers of the masseter in a tendinous raphe behind and below the mandibular angle (the pterygomasseteric sling). Because of the overall fiber orientation of the medial pterygoid muscle, it is primarily an elevator of the mandible ( Figure 42-11 ).
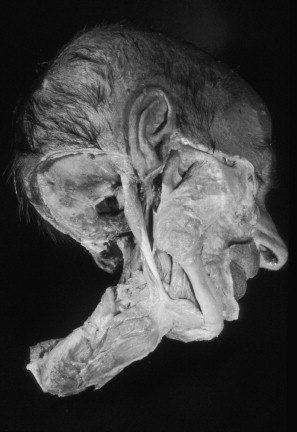
Lateral Pterygoid Muscle
The lateral pterygoid muscle arises from two heads ( Figure 42-12 ). The inferior head is about three times larger than the superior head. The superior head (sometimes called the superior pterygoid ) originates from the infratemporal surface of the greater wing of the sphenoid medial to the infratemporal crest. From its origin, the fibers of the superior head run almost horizontally backward and laterally in close relation to the external surface of the cranial base. The inferior head originates from the outer surface of the lateral pterygoid plate. Although the fibers of the inferior head also run backward and laterally, they pass upward at an angle of about 45 degrees relative to the superior head.
The two heads of the lateral pterygoid muscle are separated at their origins by a wide gap but fuse in front of the TMJ. The fibers of the superior head are attached primarily to a roughened fossa on the anteromedial surface of the condylar neck. This fossa is called the pterygoid fovea . In addition, a small portion of the superior head frequently is attached directly to the anteromedial part of the TMJ capsule and extends into the anteromedial part of the articular disc. All fibers of the inferior head insert into or along the periphery of the pterygoid fovea. This description of the lateral pterygoid is based on the work of numerous authors.
The lateral pterygoid muscle also consists of two functionally distinct parts. The superior head contracts during jaw closing, and the inferior head contracts during protraction, opening, and shifting of the jaw to one side. The superior head of the lateral pterygoid stabilizes the mandibular condyle against the articular eminence during biting and mastication. Bilateral contraction of the lower head of the lateral pterygoid muscle creates a protruding mandibular movement. Unilateral action of the inferior head shifts the midline of the mandible to the opposite side (lateral excursion). Finally, if either or both heads of the lateral pterygoid muscle are actively recruited during mandibular closure, this muscle must experience an eccentric or lengthening contraction. Under these conditions, the lateral pterygoid muscle exhibits considerable stiffness; this also facilitates joint stability by controlling or limiting condylar movements.
Digastric Muscle
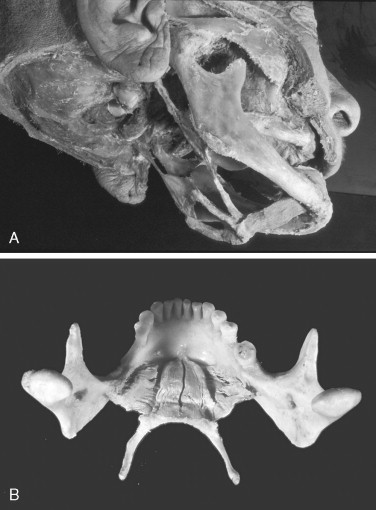
As the name implies, the digastric (two-bellied) muscle consists of an anterior belly and a posterior belly ( Figure 42-13 ). The posterior belly arises from the mastoid notch medial to the mastoid process; the intermediate tendon is held to the body of the hyoid bone by a fascial loop. The anterior belly attaches to the digastric fossa of the mandible. This fossa is located at the lower lingual border of the mandible slightly lateral to the midline. In the side view, the two bellies of the muscle form an obtuse angle. The posterior belly is much longer than the anterior belly and is only slightly flattened in the mediolateral direction.
If the hyoid is fixed by the action of the infrahyoid muscles, contraction of the digastric muscles pulls the front of the mandible back and down and thus facilitates retrusive and opening movements of the mandible. If the teeth are held firmly in occlusion (by the jaw-closing muscles), then contraction of the entire digastric elevates the hyoid as long as the infrahyoid muscles are relatively relaxed.
Mylohyoid Muscle
The mylohyoid muscle builds a muscular diaphragm or floor for most of the oral cavity (see Figure 42-13 ). It is a flat, continuous, pentagonal sheet of muscle located deep to the anterior belly of the digastric. The base of this pentagon attaches to the body of the hyoid bone, and the two adjacent sides of the base have a free edge. The remaining two sides attach to the medial or lingual aspect of the left and right mandible corpora along the mylohyoid line. The apex of this pentagon attaches to the midline lingual surface of the mandible. It also attaches to a midline mylohyoid raphe that connects the body of the hyoid with the lingual midline surface of the mandible. This raphe divides the mylohyoid into left and right sides.
The mylohyoid slightly raises the hyoid (and tongue) and floor of the mouth. Moreover, if the mandible is stabilized, it can also pull the hyoid forward. In the event that the hyoid is stabilized or is being pulled down and/or backward, this muscle also can depress the mandible.
Geniohyoid Muscle
The geniohyoid muscle is a strap-shaped muscle that runs from the ventral surface of the body of the hyoid to the lingual aspect of the mandible immediately lateral to the midline ( Figure 42-14 ). It is located superficial to the mylohyoid. The muscle is made up of nearly parallel muscle fibers that run straight from origin to insertion. The anatomy of the geniohyoid indicates that if the mandible is stabilized in occlusion, these muscles can slightly raise the hyoid (and tongue) during contraction, and it can pull these structures forward. Conversely, in the event that the hyoid is pulled downward and/or backward, the geniohyoids can depress the mandible. The geniohyoid muscles are not branchial arch derivatives. These muscles are the serial homologs of the musculi rectus abdominis; this is why they are often described as belonging to the “rectus cervicis” group—the straight muscles of the neck.
INNERVATION AND BLOOD SUPPLY
The sensory nerves of the capsule for proprioception and pain are branches of the auriculotemporal, deep temporal, and masseteric nerves. The blood supply to the capsule and disc is provided mainly by branches from a maxillary artery. Nerves ( Figure 42-15 ) and blood vessels are numerous in the posterior portions of the articular disc and fibrous capsule.
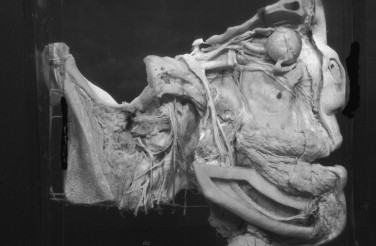
The masseter muscle is derived from the first branchial arch and therefore is innervated by the trigeminal nerve (cranial nerve V). More specifically, the masseteric nerve, a small branch of the mandibular or third division of the trigeminal nerve (V 3 ), innervates the masseter muscle. This nerve passes above the lateral pterygoid muscle and then, after passing through the mandibular notch behind the tendon of the temporalis muscle, enters the medial surface of the deep masseter muscle. The masseteric nerve supplies the deep masseter, perforates it, and then enters the superficial masseter.
The temporalis muscle is innervated by the deep temporal branches of the anterior trunk of V 3 . Of the three deep temporal nerves ordinarily present, the posterior and middle branches arise as separate filaments from the anterior trunk immediately after the trigeminal nerve emerges through the foramen ovale. The anterior branch initially is united with the buccal nerve; this common trunk, which lies in a sulcus adjacent to the foramen ovale, runs anteriorly and laterally, close to the base of the skull. It is held in place by a ligament that bridges the sulcus. If this ligament ossifies, it contributes to the formation of the temporobuccal foramen.
The anterior temporal nerve usually separates from the buccal nerve after the latter has passed between the two heads of the lateral pterygoid muscle. Its most anterior portion, however, is not positioned nearly as far forward as the anterior portion of the superficial masseter.
The nerve to the medial pterygoid arises from V 3 immediately before it divides into its anterior and posterior trunks. The medial pterygoid nerve, which also innervates the tensor tympani and tensor veli palatini muscles, reaches the medial pterygoid muscle at its upper posterior border.
The nerve to the lateral pterygoid muscle is usually a branch of the buccal nerve, which is a branch of the anterior trunk of V 3 .
The bellies of the digastric muscle are derived from the first and second branchial arches and are innervated by the mandibular division of the trigeminal (V 3 ) and facial nerves (cranial nerve VII), respectively. The mylohyoid nerve, which is a branch of the inferior alveolar nerve, innervates the anterior belly, a first-arch derivative. A branch of the facial nerve supplies the posterior belly, a second-arch derivative.
V 3 innervates the mylohyoid muscle
Stay updated, free dental videos. Join our Telegram channel
VIDEdental - Online dental courses


