http://evolve.elsevier.com/Haveles/pharmacology
The dental hygienist should become familiar with the autonomic nervous system (ANS) drugs for three reasons. First, certain ANS drugs are used in dentistry. For example, both the vasoconstrictors added to some local anesthetic solutions and the drugs used to increase salivary flow are ANS drugs. Second, some ANS drugs produce oral adverse reactions. For example, the anticholinergic drugs cause xerostomia.
Third, members of other drug groups have effects similar to those of the ANS drugs. Antidepressants and antipsychotics are drug groups with autonomic side effects, specifically anticholinergic effects. An understanding of the effects of the autonomic drugs on the body will facilitate an understanding of the action of other drug groups that have autonomic effects. Before the ANS drugs can be understood, the normal functioning of the ANS must be reviewed. The physiology of the ANS is therefore reviewed first.
Autonomic nervous system
The ANS functions largely as an automatic modulating system for many bodily functions, including the regulation of blood pressure and heart rate, gastrointestinal tract motility, salivary gland secretions, and bronchial smooth muscle. This system relies on specific neurotransmitters (chemicals that are released to send messages) and a variety of receptors to initiate functional responses in the target tissues. Before ANS pharmacology is discussed, the anatomy and physiology of this system are reviewed.
Anatomy
The ANS has two divisions, the sympathetic autonomic nervous system (SANS) and the parasympathetic autonomic nervous system (PANS). Each consists of afferent (sensory) fibers (What’s happening?), central integrating areas (Let’s coordinate all this info! Hey, what did you find out?), efferent (peripheral) motor preganglionic fibers (This is what’s happening, i.e. The heart is beating too slow), and postganglionic motor fibers (Heart, begin palpitating!).
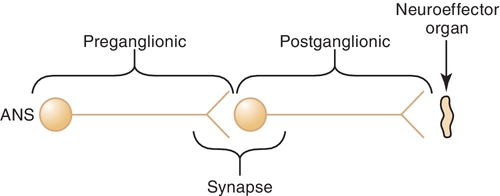
The preganglionic neuron (Figure 4-1) originates in the central nervous system (CNS) and passes out to form the ganglia at the synapse with the postganglionic neuron. The space between the preganglionic and postganglionic fibers is termed the synapse or synaptic cleft. The postganglionic neuron originates in the ganglia and innervates the effector organ or tissue.
Parasympathetic Autonomic Nervous System
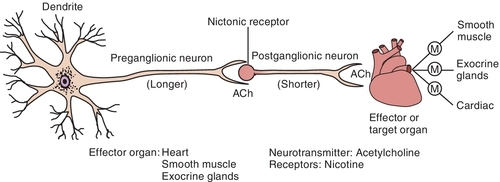
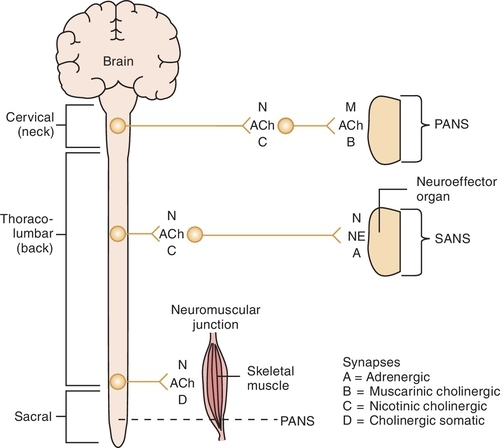
Cell bodies in the CNS give rise to the preganglionic fibers of the parasympathetic division. They originate in the nuclei of the third, seventh, ninth, and tenth cranial nerves (CN III, VII, IX, and X) and the second through the fourth sacral segments (S2 to S4) of the spinal cord. The preganglionic fibers of the PANS are relatively long and extend near to or into the innervated organ. The distribution is relatively simple for the third, seventh, and ninth cranial nerves, whereas the tenth or vagus nerve has a complex distribution. There usually is a low ratio of synaptic connections between preganglionic and postganglionic neurons, leading to a discrete response when the PANS is stimulated. The postganglionic fibers, originating in the ganglia, are usually short and terminate on the innervated tissue.
Sympathetic Autonomic Nervous System
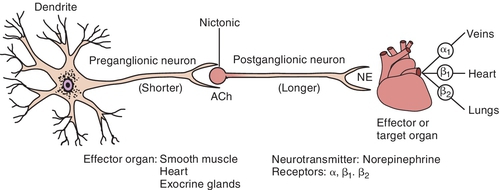
The cell bodies that give origin to the preganglionic fibers of the SANS span from the thoracic (T1) to the lumbar (L2) portion of the spinal cord (sometimes referred to as the “in-between” distribution, that is, between the two locations of the innervation of the PANS). This arrangement produces a more diffuse effect in the SANS. The preganglionic fibers exit the cord to enter the sympathetic chain located along each side of the vertebral column. Once a part of the sympathetic chain (groups of nerves a few inches from the vertebral column), preganglionic fibers form multiple synaptic connections with postganglionic cell bodies located up and down the sympathetic chain. Thus a single SANS preganglionic fiber often synapses with numerous postganglionic neurons, producing a more diffuse effect in the SANS. The postganglionic fibers then terminate at the effector organ or tissues.
The adrenal medulla is also innervated by the sympathetic preganglionic fibers. It functions much like a large sympathetic ganglion, with the glands in the medulla representing the postganglionic component. When the SANS is stimulated, the adrenal medulla releases primarily epinephrine and a small amount of norepinephrine (NE) into the systemic circulation. A diffuse response is produced when the SANS is stimulated because of the high ratio of synaptic connections between the preganglionic and postganglionic fibers and because epinephrine is released by the adrenal medulla, into the bloodstream, when stimulated.
Functional Organization
In general, the divisions of the ANS, the parasympathetic and the sympathetic, tend to act in opposite directions (Figure 4-2). The parasympathetic division of the ANS is concerned with conservation of the body processes. Both digestion and intestinal tract motility are greatly influenced by the PANS. The sympathetic division is designed to cope with sudden emergencies such as the “fright-or-flight” or “fight-or-flight” situation. In most but not all instances, the actions produced by each system are opposite: one increases the heart rate and the other decreases it; one dilates the pupils of the eye and the other constricts them. The receptors being innervated for each function may be different. For example, both the PANS and the SANS stimulate muscles in the eye that change the size of the pupil. The SANS stimulates the radial smooth muscles (out from the pupil like sun rays), producing an increase in pupil size. When the pupils are dilated the effect is termed mydriasis). The PANS stimulates the circular smooth muscles (like a bull’s-eye), producing a decrease in pupil size. When pupils are constricted the effect is termed miosis.
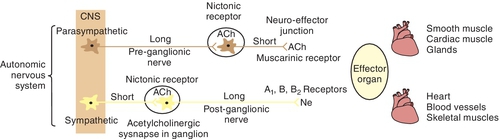
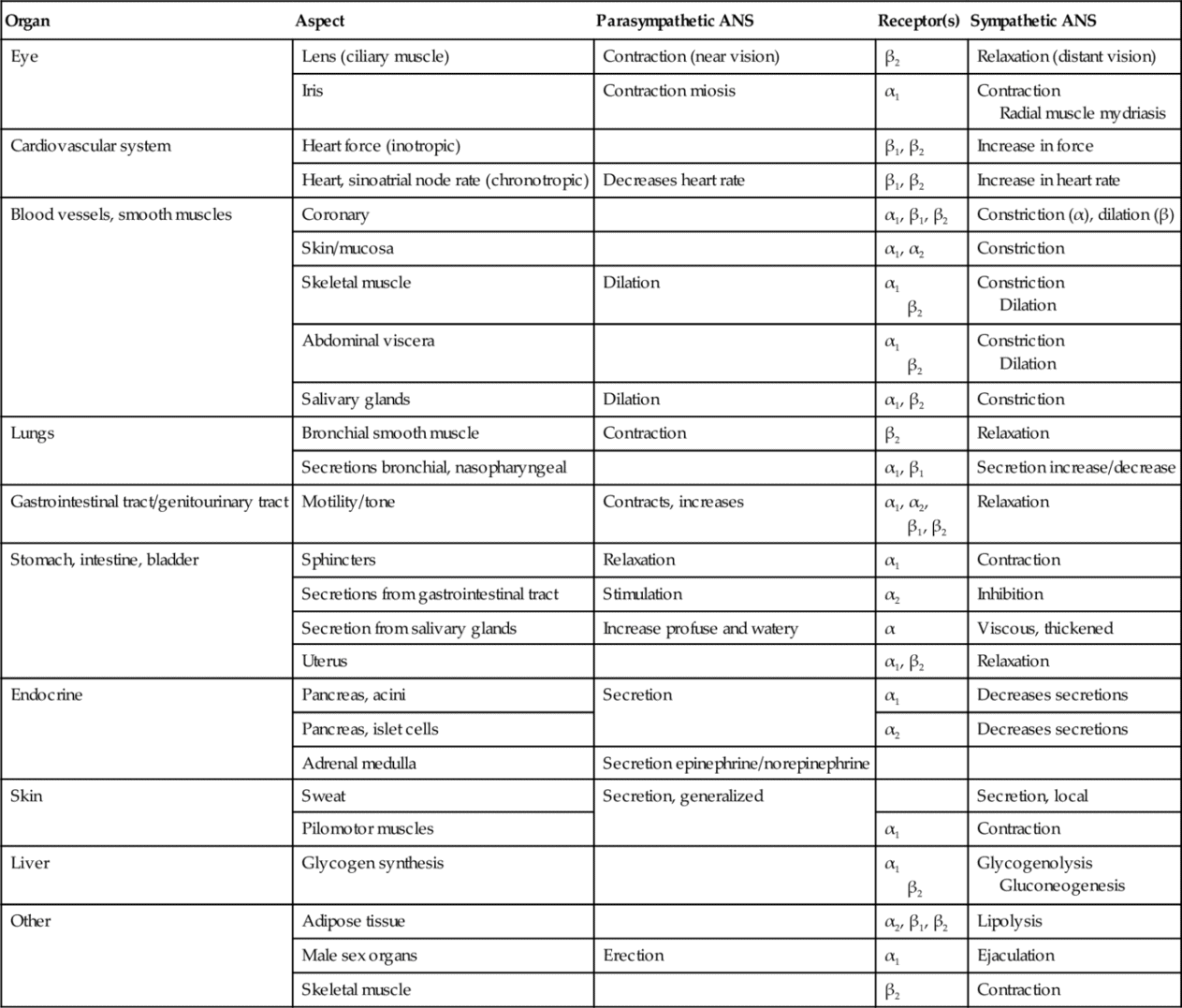
Almost all body tissues are innervated by the ANS, with many but not all organs receiving both parasympathetic and sympathetic innervation. The response of a specific tissue to stimuli at any one time will be equal to the sum of the excitatory and inhibitory influences of the two divisions of the ANS (if a tissue receives both innervations). Table 4-1 summarizes the effects of the ANS on major tissues and organ systems.
In addition to the dual innervation of tissues, there is another way in which the two divisions of the ANS can interact. Sensory fibers in one division can influence the motor fibers in the other. Thus, although in an isolated tissue preparation the stimulation of one of the divisions would produce a specific response, in the intact body a more complex and integrated response can be expected. The net effect would be a combination of the direct and indirect effects.
Neurotransmitters
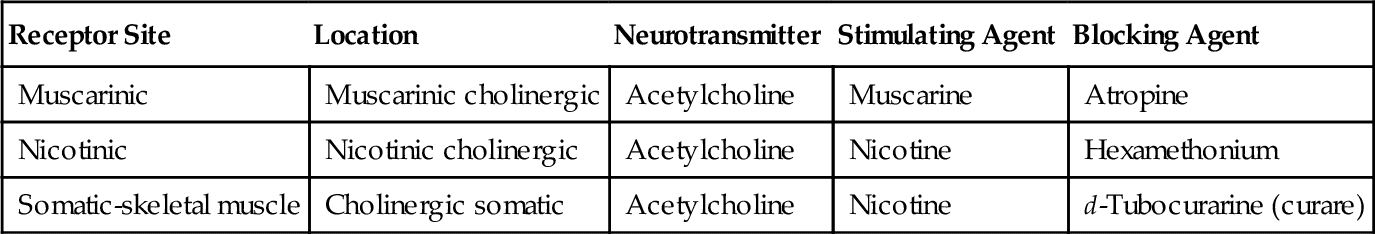
Communication between nerves or between nerves and effector tissue takes place through the release of chemical neurotransmitters across the synaptic cleft. Neurotransmitters are released in response to the nerve action potential (or pharmacologic agents in certain cases) to interact with a specific membrane component: the receptor. Receptors are usually found on the postsynaptic fiber and the effector organ but may be located on the presynaptic membrane as well (Table 4-2). The interaction between neurotransmitter and receptor is specific and is rapidly terminated by disposition of the neurotransmitter. There are several specific mechanisms by which the neurotransmitter produces an effect on the receptor.
Disposition of the neurotransmitter occurs most often by either its reuptake into the presynaptic nerve terminal or its enzymatic breakdown. Nerves in the ANS contain the necessary enzyme systems and other metabolic processes to synthesize, store, and release neurotransmitters. Thus drugs can modify ANS activity by altering any of the events associated with neurotransmitters: (1) synthesis, (2) storage, (3) release, (4) receptor interaction, and (5) disposition. The specificity of the neurotransmitters and receptors dictates the tissue response, which occurs as follows:
Parasympathetic autonomic nervous system
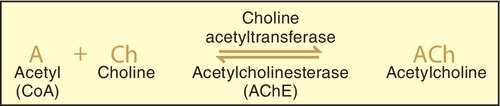
Acetylcholine has been identified as the principal mediator in the PANS. When an action potential travels along the nerve, it causes the release of the stored Ach from the synaptic storage vesicles, and if sufficient Ach is released, it initiates a response in the postsynaptic tissue. If the postsynaptic tissue is a postganglionic nerve, depolarization with generation of an action potential occurs in that neuron. In the postganglionic parasympathetic fibers, the postsynaptic tissue is an effector organ and the response is the same as that of the neurotransmitter. The action of the released Ach is terminated by hydrolysis by acetylcholinesterase to yield the inactive metabolites choline and acetic acid (or acetate) (Figure 4-7).
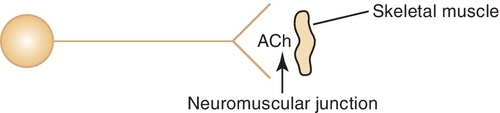
Once Ach is released from the presynaptic neuron into the synaptic cleft, it can bind to either muscarinic or nicotinic receptor sites. Small doses of Ach bind to muscarinic receptors and duplicate the effects of the chemical substance muscarine. Large doses bind to nicotinic receptors and duplicate the effects of nicotine, a chemical substance found in cigarettes. Nicotinic receptors are found in the neuromuscular junction, parasympathetic nervous system, sympathetic nervous system, adrenal medulla, and CNS, whereas muscarinic receptors are found only in the parasympathetic nervous, sympathetic nervous system, and CNS. The amount of neurotransmitter released, size of the synaptic cleft, and postganglionic site determine which receptor site is activated.
Cholinergic (Parasympathomimetic) Agents
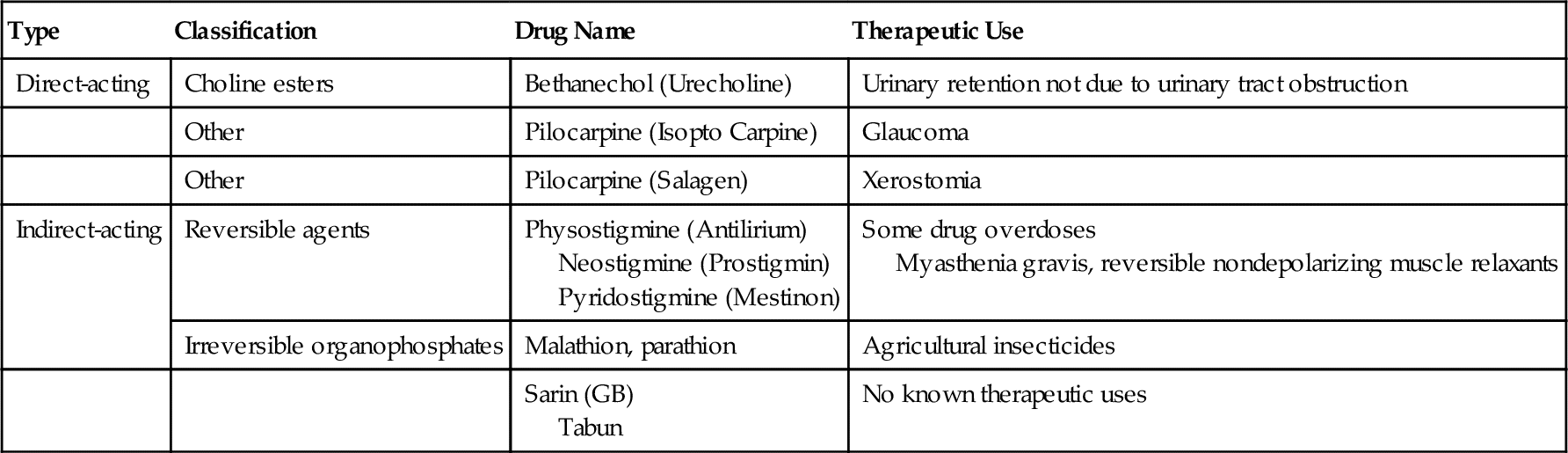
Depending on their mechanism of action (Table 4-3), the cholinergic (parasympathomimetic) agents are classified as direct-acting (acts on receptor) or indirect-acting (causes release of neurotransmitter). The direct-acting agents (Figure 4-8) include the choline derivatives and pilocarpine. The choline derivatives include both Ach and other, more stable choline derivatives. These derivatives of Ach possess activity similar to PANS stimulation but have a longer duration of action and are more selective.
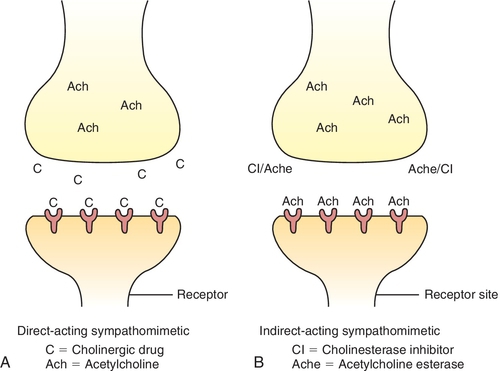
The indirect-acting (see Figure 4-8) parasympathomimetic agents or cholinesterase inhibitors act by inhibiting the enzyme cholinesterase.
When the enzyme that normally destroys Ach is inhibited, the concentration of Ach builds up (it is not being destroyed), resulting in PANS stimulation.
Pharmacologic Effects
Cardiovascular Effects
The cardiovascular effects associated with the cholinergic agents are the result of both direct and indirect actions. The direct effect on the heart produces a negative chronotropic and negative inotropic action. A decrease in cardiac output is associated with these agents.
The cholinergic agents’ effects on the smooth muscles around the blood vessels result in relaxation and vasodilation, producing a decrease in total peripheral resistance. The indirect effect of these agents is an increase in heart rate and cardiac output. Because the direct and indirect effects of these agents on the heart rate and cardiac output are opposite, the resulting effect depends on the concentration of the drug present. Generally, there is bradycardia and a decrease in blood pressure and cardiac output.
Gastrointestinal Effects
The cholinergic agents excite the smooth muscle of the gastrointestinal tract, producing an increase in activity, motility, and secretion.
Effects on the Eye
The cholinergic agents produce miosis and cause cycloplegia—a paralysis of the ciliary muscles of the eye that results in the loss of visual accommodation. Because intraocular pressure is also decreased, these agents are useful in the treatment of glaucoma.
Adverse Reactions
The adverse reactions associated with the administration of the cholinergic agents are essentially extensions of their pharmacologic effects. When large doses of these agents are ingested, the resultant toxic effects are described by the acronym SLUD: salivation, lacrimation, urination, and defecation. With even larger doses, neuromuscular paralysis can occur as a result of the effect on the neuromuscular junction. CNS effects, such as confusion, can be seen if toxic doses are administered.
The treatment of an overdose of cholinesterase inhibitors, such as the insecticides or organophosphates (parathion), includes a combination of pralidoxime (pra-li-DOX-eem) (2-PAM, Protopam) and atropine. Pralidoxime regenerates the irreversibly bound Ach receptor sites that are bound by the inhibitors (knocks them off like a prizefighter), and atropine blocks (competitively) the muscarinic effects of the excess Ach present.
Contraindications
The relative contraindications to or cautions with the use of the cholinergic agents stem from these agents’ pharmacologic effects and adverse reactions. They include the following:
Uses
The direct-acting agents are used primarily in the treatment of glaucoma, a condition in which the intraocular pressure is elevated. Occasionally, they are used to treat myasthenia gravis, a disease resulting in muscle weakness from an autoimmune reaction that reduces the effect of Ach on the voluntary muscles. The urinary retention that occurs after surgery is also treated with the choline esters (see Table 4-3).
Pilocarpine (pye-loe-KAR-peen) (Salagen), a naturally occurring cholinergic agent, is used in the treatment of xerostomia, but its success may be limited because of the myriad of potential side effects. Common side effects from pilocarpine include perspiration (sweating), nausea, rhinitis, chills, and flushing. Pilocarpine is available in 5-mg tablets. The usual dose of pilocarpine is 5 mg three times a day (tid). This can be obtained by giving one 5-mg tablet tid [three times a day]). Pilocarpine is also available as ophthalmic solution in strengths ranging from 0.5% to 10%. It is used topically in the eye to treat glaucoma. Several strengths (e.g., 2%) are available as generic preparations.
The indirect-acting cholinergic agents, the cholinesterase inhibitors, are divided into groups on the basis of the speed of reversibility of their binding to the enzyme. Edrophonium is rapidly reversible, whereas physostigmine and neostigmine are slowly reversible. These agents are used to treat glaucoma and myasthenia gravis.
Stay updated, free dental videos. Join our Telegram channel
VIDEdental - Online dental courses


