Chapter 1
Basic Biology of Oral Microbes
Abstract
This chapter describes the basic biology of oral microbes, including cytological basis of the microorganisms, microbial morphology, microbial cell structure, microbial physiology, and microbial genetics.
Keywords
Bacteria; Cell wall; Cytoplasm; Fungi; Heredity; Physiology; Transduction; Variation; Virus
1.1. Cytological Basis Of Microorganisms
The cell is the fundamental unit of all living organisms. Various subunit structures and chemical substances found on and inside the cell make complex cellular functions possible. Microbes can be divided into three major groups according to their morphological structure, degree of differentiation, and chemical composition: eukaryote, prokaryote, and acellular microorganisms (Figure 1.1(A)–(D)).
1.2. Microbial Morphology
Microorganisms, also known as microbes, are tiny organisms that are only visible under an optical microscope or an electron microscope. They are small in size and simple in structure. Microbes reproduce quickly, can tolerate a wide range of environmental conditions, are widely distributed, highly variable, and tend to congregate.
1.2.1. Microbial Size
As many types of microbes exist, they vary widely in size. Generally, the units used to measure microbes are μm and nm. Most cocci are 1 μm in diameter. Bacilli can be further divided into coccobacilli, brevibacteria, and long bacilli, and measure approximately 1–10 μm in length and 0.3–1 μm in width. Spirochetes measure approximately 6–20 μm in length and 0.1–0.2 μm in width. Fungi are several times larger than bacteria. Most viruses are smaller than 150 nm and are only visible under the electron microscope. The same microbes can change in size depending on their environment or age (Figure 1.2).
1.2.2. Microbial Morphology
Different types of microbes have different, but characteristic, shapes. Under suitable conditions, the shape and size of microbes are relatively stable. It is important to know the morphological structure of microbes, as it provides us with a better understanding of microbial physiology, pathogenic mechanisms, antigenic features, and allows us to identify them by species. In addition, knowledge of microbial morphology can be helpful in diagnosing disease and in preventing microbial infections.
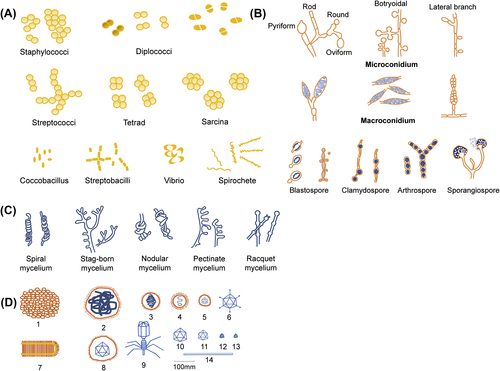
1. Bacteria are complex and highly variable microbes. They come in four basic shapes: spherical (cocci), rod-shaped (bacilli), arc-shaped (vibrio), and spiral (spirochete) (Figure 1.3(A)).
2. Fungi are divided into unicellular and multicellular according to the number of cells that make up the organism. Unicellular fungi, such as Saccharomyces and other yeast-like fungi, are usually round or oval. Multicellular fungi have hyphae and spores. The hyphae and spores of different fungi are shaped differently (Figure 1.3(B) and (C)).
3. Many viruses are spherical or almost spherical, some are rod-shaped (often seen in plant viruses), filamentous (e.g., freshly isolated influenza virus), bullet-shaped (e.g., rabies virus), brick-shaped (e.g., poxvirus), and tadpole-shaped (e.g., bacteriophage) (Figure 1.3(D)).
1.3. Microbial Cell Structure
Although different microbes possess different cellular structures, there are certain commonalities within groups of microbes.
1.3.1. Basic Bacterial Structures
The architecture of bacterial cells consists of basic and special structures. Basic structures include the cell wall, cell membrane, cytoplasm, nuclear material, ribosome, plasmid, etc. Special structures, which are only found in some bacteria, include the flagellum, pilus, capsule, spore, etc. (Figure 1.4).
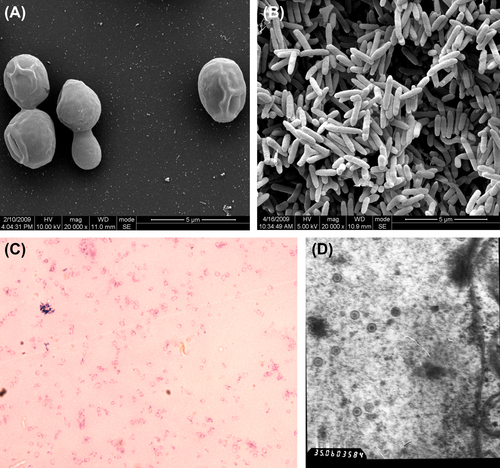
Figure 1.1 (A) Eukaryotic microbes (Saccharomyces, SEM): The eukaryotic cell has a high degree of nuclear differentiation. It has a nuclear membrane, nucleoli, and chromosomes. There is a complete complement of structured organelles in the cytoplasm and cellular reproduction takes place by mitosis. Examples include fungi and algae. (B) Prokaryotic microbes (bacteria). (C) Prokaryotic microbes (mycoplasma): The prokaryotic cell has a primitive nucleoplasm and cell membrane; it has no nuclear membrane, nucleolus, or organelles. Prokaryotes include bacteria such as Mycoplasma, Chlamydia, and Rickettsia. (D) Acellular microorganisms (herpes simplex virus): Acellular microorganisms are the smallest microorganism with no typical cell structure and no enzymatic energy-production system. They consist merely of a nucleic acid genome (DNA/RNA) and a protein coat (the capsid). Acellular microorganisms can only reproduce inside a living cell. Examples include viruses and subviral agents.
1.3.1.1. Cell Wall
The cell wall is the outermost structure of the bacterial cell and is located outside the cell membrane. It is transparent, tough, and flexible. The average thickness ranges from 15 to 30 nm. It mainly consists of peptidoglycan, also called mucopeptide, glycopeptide, or murein. Bacteria are classified into gram-positive and gram-negative based on the appearance of the cells after Gram stain. The peptidoglycan of gram-positive bacteria is composed of a glycan backbone, tetrapeptide side chains, and a pentapeptide cross-linking bridge (Figure 1.5(A)). The peptidoglycan of gram-negative bacteria is composed of a glycan backbone and a tetrapeptide side chain (Figure 1.5(B)).
Gram-positive and gram-negative bacteria have unique structures other than peptidoglycan in their cell walls (Figure 1.5(C) and (D)). Other substances, such as compound polysaccharide, surface protein, proteins M and G of Streptococcus, protein A of Staphylococcus aureus, etc. are found on the outer layer of the cell wall of some gram-positive bacteria.
1.3.1.2. Cell Wall-Deficient Bacteria (Bacterial L Form)
Cell wall-deficient bacteria are strains of bacteria that lack cell walls. The peptidoglycan that makes up the cell wall can be destroyed or inhibited by physical, chemical, or biological factors. When gram-positive bacteria lack a cell wall, the cytoplasm is surrounded by the cell membrane, and the entire structure is known as a protoplast. When gram-negative bacteria do not have a cell wall, the cytoplasm is protected by the outer membrane, and the entire structure is called a spheroplast. Bacteria that have lost their cell wall are still capable of growing and dividing as cell wall-deficient bacteria. Examples of these were first isolated in 1935 by Emmy Klieneberger-Nobel, who named them “L-forms” after the Lister Institute in London where she was working at the time. L-form bacteria give rise to a variety of cell morphologies and sizes and can be spherical, rod-shaped, filiform, etc. The rate of growth and division of L-form bacteria is slow. They also form distinctive bacterial colonies when plated on agar. Some L-form strains have a tendency to revert to the normal phenotype when the conditions that were used to produce the cell wall deficiency are reduced. L-form bacteria are difficult to stain or stain unevenly. In a Gram stain test, L-form bacteria always show up as gram-negative, due to the lack of a cell wall.
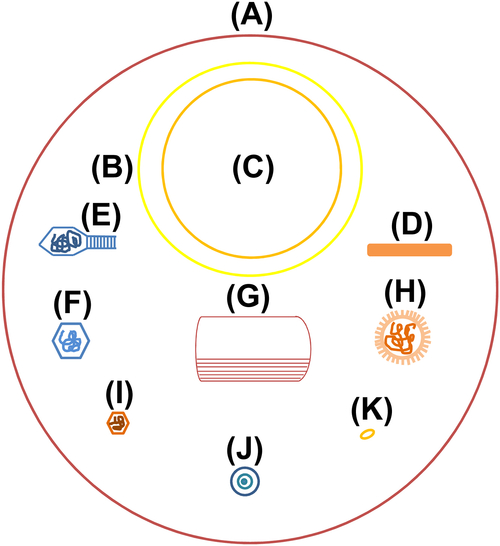
Figure 1.2 Microbial size
(A) Staphylococcus 1000 nm. (B) Rickettsia 450 nm. (C) Chlamydia 390 nm. (D) Tobacco mosaic virus 300 × 15 nm. (E) Escherichia coli bacteriophages, head 65 × 95 nm, tail 12 × 100 nm. (F) Adenovirus 70 nm. (G) Poxvirus 300 × 230 nm. (H) Influenza virus 100 nm. (I) Poliovirus 30 nm. (J) Japanese encephalitis virus 40 nm. (K) Molecule of egg protein 10 nm.
1.3.1.3. Cell Membrane
The cell membrane is a selectively permeable biological membrane found inside the cell wall and surrounding the cytoplasm. It is made of a lipid bilayer. The cell membrane is compact and flexible, and measures approximately 7.5 nm in thickness. It accounts for 10–30% of the bacterial cell dry weight. The structure of the bacterial cell membrane resembles that of eukaryotic cell membranes, except it is deficient in cholesterol. The lipid bilayer is embedded with carrier proteins and zymoprotein, which possess specific functions.
The cell membrane of some bacteria can form invaginations into the cytoplasm called mesosomes.
1.3.1.4. Cytoplasm
The cytoplasm is the gel-like substance enclosed within the cell membrane, which is made up of water, proteins, lipids, nucleic acids, inorganic salts, etc. Most metabolic activities take place within the cytoplasm, and subcellular structures, such as ribosomes, plasmids, and cytoplasmic granules, are located in the cytoplasm.
Ribosomes are found in cytoplasm. They are approximately 15–20 nm in diameter and are composed of a small (30S) and a large (50S) subunit. The association between subunits requires the presence of Mg2+. Ribosomes are made up of 30% ribosomal proteins and 70% ribosomal RNA.
Plasmids are small, circular, double-stranded DNA molecules and are extrachromosomal genetic material. They can replicate independently of chromosomal DNA and transmit genes encoding drug resistance, bacteriocins, toxins, and more from one bacterium to another via conjugation and transduction.
Cytoplasmic granules is a general term referring to many types of cytoplasmic inclusion granules. They are an intracytoplasmic (inside the cytoplasm of a cell) form of storing nutrients and energy and include molecules such as polysaccharides, lipids, phosphates, etc. They are not essential or permanent structures in cells. Cytoplasmic granules are also known as metachromatic granules because they may stain into different colors than other bacterial cell structures.
1.3.1.5. Nuclear Material
The bacterial nuclear material is also called the nucleoid. It is a piece of double-stranded DNA devoid of nuclear membrane, nucleolus, or histones and is the bacterial equivalent of chromatin. The function of the nucleoid is similar to that of the nucleus in eukaryotic cells and encodes genes necessary for activities and traits such as growth, metabolism, reproduction, heredity, mutation, etc.
1.3.2. Special Bacterial Structures
1.3.2.1. Capsule
The capsule is a layer of slime that lies outside the bacterial cell wall. It is secreted by bacteria and diffuses into the surrounding medium. Based on its appearance when examined by light microscope, the bacterial capsule is classified into two types: microcapsule, which is less than 0.2 μm in thickness and escapes optical detection; and capsule or large capsule, which is over 0.2 μm in thickness, binds tightly to the cell wall, and presents an obvious boundary under optical microscope. The capsule shows up as negatively stained when ordinary staining techniques are used. It appears as a clear halo around the bacterium when stained samples are examined by light microscope. Using special staining, the capsule can be stained differently from the bacterial cell (Figure 1.6). Most bacterial capsules are composed of polysaccharides, but a few capsules are made of polypeptides.
Stay updated, free dental videos. Join our Telegram channel
VIDEdental - Online dental courses


