4
Dental caries and pulpitis
- The aetiology, epidemiology, microbiology and development of the carious process
- The carious lesion and some of the important management rationales
- The operative techniques and technologies used to treat dental caries
Introduction
Dental caries is the major cause for loss of teeth in children and adults. It affects the enamel surfaces of teeth in children and adults and the exposed root surfaces of the teeth of the elderly. Dental caries is the result of the interaction of the bacteria on the tooth surface, the dental plaque or oral biofilm, the diet and specifically fermentable carbohydrate components of the diet, which are fermented by the plaque microflora to organic acids (primarily lactic and acetic acids), and the teeth acting together over time (Figure 4.1). Dental caries will occur only when the three factors are present together and when they act together over a sufficient period of time.
Figure 4.1 Schematic diagram to demonstrate the requirement for diet, bacteria and dentition to interact together over time to initiate caries.
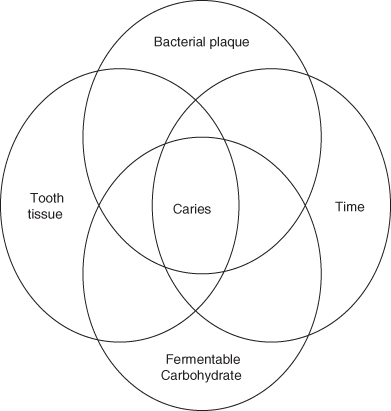
If the different factors are considered individually, it is possible to obtain a better understanding of the nature of their interactions and how these interactions may manifest as a carious lesion and, eventually, if not treated either preventatively or with a restoration, as pain and ultimately in the loss of the tooth.
Aetiology and microbiology of dental caries in relation to dental plaque
Dental plaque is a natural biofilm (Figure 4.2). The number of different bacterial species in dental plaque is not known, but sophisticated molecular biology studies have shown that over 700 different species may be identified; many more have yet to be characterised. The majority of these species have not been cultured in the laboratory using the current methods for bacterial isolation. It is also worthwhile noting that these molecular approaches have demonstrated that, for example, Streptococcus mutans, previously reported only to be isolated from the oral cavity after the eruption of the lateral incisors, is actually present in the mouth, colonising the tongue, within a few months of birth.
Figure 4.2 Anterior view of the dentition with 36 hour accumulation of plaque. (Reproduced with permission of Oxford University Press, Banerjee A. Watson TF (2011) Pickard’s Manual of Operative Dentistry, 9th edition.)
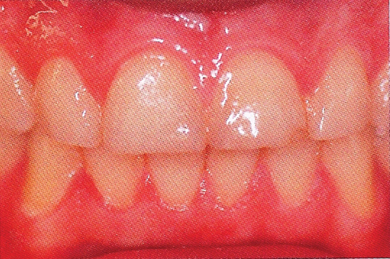
A biofilm is a community of microorganisms attached to a solid surface, with the bacteria encapsulated in polymers derived from the bacteria and exhibiting specific biofilm characteristics, including increased resistance to antimicrobials and biocides (chemicals used to kill bacteria) and the production of novel proteins. An appreciation of this type of growth has led to attempts to develop novel antiplaque strategies, which target the inhibition of function of novel biofilm-specific proteins.
Dental plaque biofilm is present on virtually all tooth surfaces and has a general structure whereby salivary and some bacterial proteins are initially deposited on the enamel surface to form the pellicle and the pellicle then absorbs further bacteria. The first bacteria to bind to the tooth surface, the pioneer organisms, include Streptococcus oralis, Streptococcus sanguinius, Neisseria and Haemophilus species and Actinomyces naeslundii. These bacteria are isolated from cleaned tooth surfaces within 60 minutes of cleaning. Following the binding of these bacteria, other organisms bind and eventually plaque of full thickness develops, partly because of bacterial binding but primarily because of bacterial growth. The binding of bacteria to the pellicle and to other bacteria is mediated by specific interactions in which only certain bacteria bind to components of the pellicle and only certain bacteria interact. One of the major and most interactive of all the bacteria in dental plaque is Fusobacterium nucleatum which binds to many other bacteria; it acts almost as a nucleus for plaque formation, cross-linking otherwise non-interacting bacterial species (Figure 4.3).
Figure 4.3 Arrangements of bacteria within dental plaque showing potential attachments between bacteria and components of the pellicle (Reproduced from Kolenbrander, P.E., Andersen, R.N., Blehert, D.S., Egland, P.G., Foster, J.S. and Palmer, R.J. Jr. (2002) Communication among oral bacteria. Microbiology and Molecular Biology Reviews, 66(3), 486–505, with permission from ASM Press.)
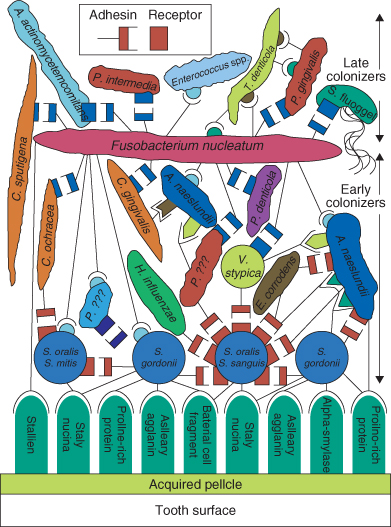
The composition of dental plaque varies over time. In supragingival plaque the flora becomes more anaerobic, with pigmented anaerobic rods such as Prevotella and Fusobacterium species becoming increasingly abundant as the plaque accumulates and ages, as it becomes a climax community. However, in sites where caries is initiated, in the stagnation sites (pits and fissures on the occlusal surfaces and at approximal sites on the enamel and at the gingival margin on exposed root surfaces) the composition of the plaque biofilm reflects the local acidic environment. Organisms most likely to be isolated from these stagnation sites are Streptococcus mutans and Streptococcus sobrinus (mutans streptococci), other streptococci (S. oralis, S. intermedius and S. anginosus), lactobacilli, Actinomyces and Bifidobacterium species. These organisms are Gram-positive, acidogenic (producing acid from sugars) and each includes strains which are aciduric (able to survive in acidic environments). At these stagnation sites yeasts, such as Candida albicans, may also be present as they are also both acidogenic and aciduric. Veillonella species, which are Gram-negative cocci, are also isolated, as this genus has the ability to use lactic acid produced from these other species to generate energy for growth.
Although it has long been known that bacteria are present on the teeth it was not until the 1950s that it was proven that without bacteria there was no tooth decay. Only in recent years has it become clear that the identification of the causative microbiological agents of tooth decay remains elusive.
Lactobacilli were regarded as the main microbiological agent of dental caries for many years until the 1940s, as they were, and still are, isolated in great numbers from active carious lesions. However, a definitive longitudinal study clearly demonstrated that lactobacilli colonised lesions after their formation and that they were not prevalent in dental plaque during the period of lesion formation (Hemmens et al., 1946). Since lactobacilli proliferate in acidic environments, they are good indicators of the amount of sugar that patients consume. Monitoring the levels of these bacteria in saliva is a good way of establishing whether patients are following dietary advice and using adequate oral hygiene measures.
The studies which established once and for all that bacteria were essential for the initiation of tooth decay were carried out in the early 1950s using germ-free (gnotobiotic) rats. In these studies, rats were inoculated orally with single strains of bacteria isolated from the faeces of rats fed sucrose-rich diets and with tooth decay. Only rats inoculated with bacteria and fed sucrose in the diets developed caries. However, not all bacterial isolates were capable of forming carious lesions. In addition, bacteria, including Streptococcus salivarius and Enterococcus spp., which are not normally isolated in large numbers from dental plaque, also caused caries in rats. From these studies, Streptococcus mutans, isolated many years previously, appeared to be the most cariogenic of all the species tested, while other commensal bacteria (e.g. streptococci and Actinomyces), were less so. Some strains of Streptococcus oralis and milleri-group streptococci produced levels of caries that approached those found with Streptococcus mutans and Streptococcus sobrinus. However, all strains of mutans streptococci produced high levels of caries while only a few of the many other species, especially other species of streptococci, did so. It is now apparent that these non-mutans streptococci are heterogeneous (different) with respect to their ability to cause caries and that this is reflected in the range of acidogenic and aciduric properties of these organisms compared to the relatively homogeneous (similar) properties of the mutans streptococci.
It was for these reasons that for many years, mutans streptococci were regarded as the bacteria associated with the initiation of dental caries and the levels of these organisms were used to assess caries risk. Consequently, considerable research was undertaken to establish whether Streptococcus mutans was the cause of caries in humans. Despite definitive evidence that this was the case, a number of investigations were undertaken attempting to develop novel treatments to eradicate S. mutans from the oral cavity as a means of inhibiting caries formation. The methods which have been investigated include:
- Vaccine development.
- Passive immunisation with the direct application of anti-S. mutans antibodies directly on the tooth surface.
- Replacement therapies in which less cariogenic bacteria are introduced into the mouth to eradicate the more cariogenic S. mutans strains.
Some of these approaches work in laboratory animal models, and some eradicate S. mutans for short periods from the human oral cavity, but none has yet been shown to be effective at reducing the incidence of dental caries in humans. The reasons for the lack of success are still being determined. It is, however, well established that there is a clear association between the numbers and concentrations of S. mutans in plaque and/or saliva and the dental caries status of populations, and to a lesser extent to the caries status of an individual. It is also known that dental caries occurs in the absence of S. mutans and individuals with high levels of S. mutans do not necessarily have to have caries.
Caries-associated characteristics of caries-associated microorganisms
Laboratory investigations into the characteristics of bacteria, which might explain their involvement in the caries process, have focused on those related to interactions with sucrose, as it is the most cariogenic sugar and many bacteria make sticky polymers from it which may facilitate the adhesion of bacteria to the tooth surface and facilitate the caries process. As caries is mediated by acids formed by bacteria, the ability to produce large amounts of acid (acidogenicity) rapidly, to tolerate exposure to low pH environments (aciduricity) and to produce acid in an already acidic environment were considered features that bacteria initiating dental caries should exhibit. It was soon established that, in comparison with other bacteria, mutans streptococci are generally able to produce acid more rapidly, to produce the most acid in a low pH environment (pH 5.5 or less) and to survive exposure to acidic conditions. The mechanisms underlying the aciduricity of S. mutans have been investigated using modern molecular and protein analysis methods, but to date no precise mechanism has been identified. It is unlikely that the expression of any single gene is responsible solely for the aciduricity of mutans streptococci.
Mutans streptococci also possess a number of extracellular enzymes (glucosyltransferases and fructosyltransferase), which form a range of distinct polymers from sucrose. Uniquely, mutans streptococci produce a highly branched, water-insoluble glucan called mutan, which may facilitate its establishment in dental plaque. These polymers are necessary for the adherence of S. mutans to enamel; in rat studies, mutants with these genes inactivated and unable to produce these polymers are less able to colonise the dentition of rats and consequently are less able to initiate carious lesion formation. These characteristics have been extensively studied in mutans streptococci, but evidence has recently emerged that bacteria other than mutans streptococci also have these same acidogenic and aciduric properties and produce extracellular polymers promoting plaque formation.
It is difficult to find relevant information on the cariogenic characteristics of other oral bacteria other than the mutans streptococci since so much emphasis has been placed on enumerating mutans streptococci in dental plaque and analysing their physiological properties. However, a study of elderly Chinese subjects related the sucrose-induced pH response of dental plaque on sound exposed root surfaces to the microbial composition of the overlying plaque (Aamdal-Scheie et al., 1996). The prevalence of a range of bacteria, including mutans streptococci, was determined and the pH of the plaque measured before and after the application of a sucrose mouth rinse. The plaque pH response on sound and carious root surfaces to sucrose was the same regardless of the presence or absence of mutans streptococci (Figure 4.4). Clearly, the rate of acid production from dental plaque was not determined by the presence of mutans streptococci nor was the final pH recorded following the sucrose rinse determined by the presence of mutans streptococci in the oral biofilm.
Figure 4.4 Stephan curves showing pH change in plaque of subjects with and without Streptococcus mutans (Reproduced with permission from Beighton, D. (2005) The complex oral microflora of high-risk individuals and groups and its role in the caries process. Community Dentistry and Oral Epidemiology, 33, 248–255. John Wiley & Sons, Ltd.)
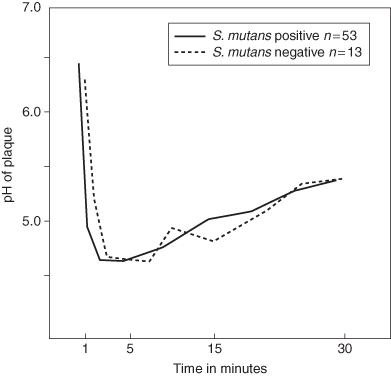
These in-vivo observations are different to those derived from in-vitro laboratory studies using single strains of planktonic organisms (organisms growing in liquid culture). The bacteria on the teeth grow in a biofilm and they grow as part of a very complex interacting microbial community. None of these features are present in many of the in-vitro studies, which found the acidogenicity of mutans streptococci to be significantly greater than that of other bacteria isolated from dental plaque. The results from the Chinese subjects clearly show that the magnitude of the cariogenic challenge was not determined by the presence of mutans streptococci and that other bacteria present in the dental plaque were responsible for caries initiation and progression. This idea that bacteria other than mutans streptococci are associated with the initiation and progression of dental caries is now well accepted and new data have suggested roles for many other bacterial species in the caries process.
The acidogenicity and aciduricity of plaque bacteria and evidence for a multispecies aetiology of dental caries
The acidogenic composition of dental plaque reflects the dietary habits and caries status of an individual. A high acidogenic bacterial level was associated with active lesions or recent restorations, while individuals who were caries-free had a lower level of acidogenic bacteria. The acidogenic bacterial population also increased as the frequency exposure to dietary sugars increased. Streptococci, Actinomyces and S. mutans are also isolated amongst the acidogenic flora from dental plaque but no consistent relationship was found between either their isolation frequency or proportions and caries status, suggesting that the acidogenic population in plaque was heterogeneous. It was also apparent that bacteria other than mutans streptococci increased in plaque in response to an increased frequency of consumption of sugars.
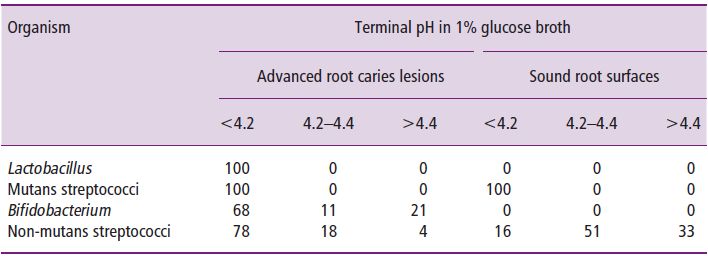
It has been the studies of van Houte and colleagues (van Houte et al., 1996) into the acidogenicity of bacteria isolated from a variety of sound and carious tooth sites that has shed most light on the role of bacteria other than mutans streptococci in the caries process. They found that other bacteria in dental plaque, presumably including S. gordonii, S. oralis, S. mitis and S. anginosus, outnumber the mutans streptococci in virtually all plaque samples. It was a logical extension of these observations to conclude that these bacteria may play a significant role in the caries process. There was clearly a selection for more acidogenic strains of non-mutans streptococci within carious lesions compared to those isolated from plaque on sound tooth surfaces (Table 4.1). It can be seen that lactobacilli and mutans streptococci all produced a terminal pH that was <4.2 while the other bacteria were heterogeneous. Of 100 isolates from root lesions, 78 produced a final pH of <4.2 but only 16 of 100 isolates from sound root surfaces produced the same low pH. van Houte and colleagues (1996) have reported that the emergence of mutans streptococci in plaque was usually preceded by an increase in the numbers of other types of acidogenic bacteria, including non-mutans streptococci. It was suggested that the initiation of caries, in the absence or presence of mutans streptococci, could be explained by the dynamic and positive relationship among the following factors:
Table 4.1 Comparison of the percentage distribution of lactobacilli, mutans streptococci, Bifidobacterium and non-mutans streptococci, isolated from root caries lesions (n = 84) and sound tooth surfaces (n = 223) on the basis of their terminal pH in glucose broth. (Reproduced with permission from van Houte, J., Lopman, J. and Kent, R. (1996) The final pH of bacteria comprising the predominant flora on sound and carious human root and enamel surfaces. Journal of Dental Research, 75(4), 1008–1014.)
- Carbohydrate consumption.
- Plaque flora composition.
- Plaque acidogenic potential.
- Caries activity.
The studies by van Houte reveal a little of the as yet poorly understood relationships between diet, microflora and caries initiation. However, they clearly demonstrate the heterogeneity of oral bacteria which may be exacerbated by exposure to fermentable dietary carbohydrates, and the consequent increase in acid production, by in turn exhibiting increased acidogenicity.
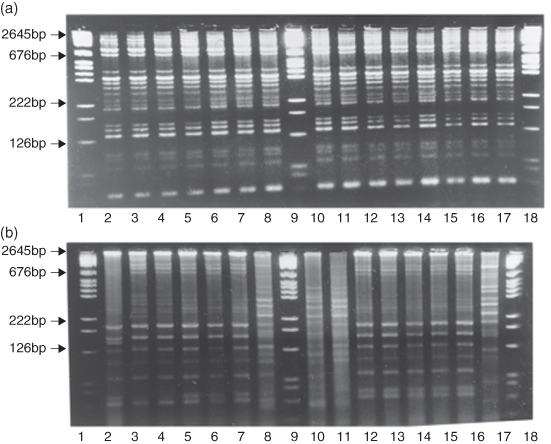
The aciduric flora of dental plaque has not been extensively studied in relation to dental caries initiation or progress. The variability observed by van Houte (1996) in respect to acidogenicity has also been observed for the aciduricity of dental plaque bacteria using molecular biology techniques (Figure 4.5). Here S. oralis strains were isolated from different people on media at pH 5.2 (acidic) and pH 7.0, and they were DNA fingerprinted using a method called repetitive extragenic palindromic polymerase chain reaction (REPPCR). These studies showed that this species was heterogeneous and that there exist distinct aciduric populations of S. oralis. Using the same methods the aciduric bacteria associated with root caries in the elderly were investigated by culturing plaque from different root surfaces on media at acid pH values. Plaque was taken from sound exposed root surfaces in subjects with no root caries activity, from sound root surfaces in subjects with root caries and from root carious lesions. It was found that the proportion of the flora that was isolated at pH 4.8 was significantly less from the sound surfaces in subjects with no root caries (0.008%) compared to the levels in the other groups, which were not significantly different. At pH 5.2 the recovery from the lesions was significantly greater than from the other two sites. From none of these samples were mutans streptococci recovered amongst the predominant aciduric bacteria. This study shows that the most diseased sites and those most at risk harboured the largest populations of aciduric bacteria, while the sites in subjects at least risk harboured the smallest populations of aciduric bacteria and that very few of the acidogenic bacteria isolated from these sites were mutans streptococci.
Figure 4.5 DNA fingerprint patterns of Streptococcus oralis. Strains grown at pH 7.0 or pH 5.2 from the same plaque sample. Evidence of heterogeneity within a species related to strain aciduricity. (Reproduced from Alam, S., Brailsford, S., Adams, R.S., Allison, C., Sheehy, E., Zoitopoulos, L., Kidd, E.A. and Beighton, D. (2000) Genotypic heterogeneity of Streptococcus oralis and distinct aciduric subpopulations in human dental plaque. Applied and Environmental Microbiology, 66, 3330–3336, with permission from ASM Press.)
Together, these studies, derived using essentially conventional culture techniques supplemented with an assessment of genotypic and physiological characteristics, provide evidence that the microflora is heterogeneous. Individual species may be heterogeneous with respect to acidogenicity and aciduricity, features of mutans streptococci used to implicate these species in the caries process. While the associations between caries and the proportions of bacteria with acidogenic or aciduric traits were demonstrated, these studies provide only a basis for designing better investigations to enable a deeper understanding of the involvement of bacteria in the process of lesion formation. The changes in the flora from sound surface to white spot lesion through to cavitated lesion are not at all understood. The cross-sectional studies described clearly suggest microbial succession between these stages, but a definitive description of these flora and their physiological characteristics needs to be determined.
When it comes to considering the whole of the microflora of the dental plaque associated with dental caries, the newer techniques for studying bacterial populations provide a completely different insight into the causative bacterial species than has been previously understood from studies designed to associate mutans streptococci with caries. The modern studies use non-cultural methods and involve extracting DNA from whole plaque and analysing this to determine the number of species present or looking for the presence of a wide range of different, pre-determined bacteria.
From a study by Becker and colleagues (Becker et al., 2002), the 16s rRNA genes, which are different in each bacterial species, were identified from dental plaque associated with sound and carious coronal sites in children. They analysed a total of 294 genes and found that these belonged to 68 species or phylotypes, of which 18 were uncultivated taxa and 10 were not previously identified; so 28 of 68 had not been previously encountered. The most complex flora was associated with plaque from sound intact tooth surfaces. Similar observations were made by Wade and Banerjee (Munson et al., 2004) who investigated the microflora of five carious teeth and identified 95 different bacterial species amongst their samples, of which 44 were detected by the molecular method alone; 31 had never been previously described. All these studies demonstrate the great diversity of the microflora associated with dental plaque in both health and disease.
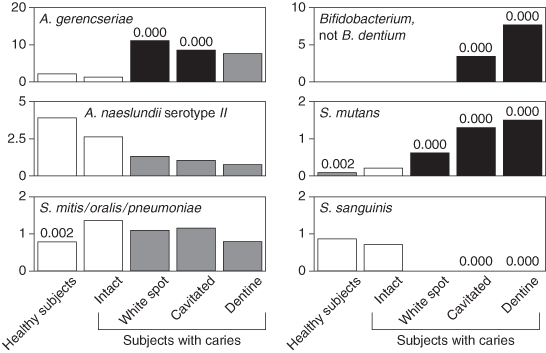
Using a more targeted approach, investigators at Harvard University used a semiquantitative method to determine the relationships between the presence and numbers of 23 well characterised species and the health or dental caries experience in children. They found the often-reported strong relationship between the numbers of S. mutans and caries but not between S. sobrinus and caries. However, other relationships were also demonstrated and these were that Actinomyces gerensceriae, Bifidobacterium, Veillonella, S. salivarius, S. constellatus, S. parasanguis and Lactobacillus fermentum were also associated with caries. A. gerensceriae and other Actinomyces spp. appeared to be associated with caries initiation and a new Bifidobacterium species was associated with deep caries lesions (Figure 4.6).
Figure 4.6 Relationship between bacterial numbers and caries status of teeth in children. (Reproduced from Becker, M.R., Paster, B.J., Leys, E.J., Moeschberger, M.L., Kenyon, S.G., Galvin, J.L., Boches, S.K., Dewhirst, F.E. and Griffen, A.L. (2002) Molecular analysis of bacterial species associated with childhood caries. Journal of Clinical Microbiology, 40(3), 1001–1009, with permission from ASM Press.)
These studies only serve to underline the complexity of the flora associated with caries and with caries initiation. They demonstrate that the proposed relationship between the presence of mutans streptococci and the aetiology of dental caries may be facile and simplistic. No matter how useful the relationship between the salivary or plaque levels of mutans streptococci and caries is in clinical practice, it must be regarded as an association and not a cause and effect relationship (see below). Caries occurs due to a complex interaction between dental plaque, whose physiological characteristics may be modified by carbohydrates in the diet, the diet itself and oral hygiene practices. These interactions are difficult to unpick and understand, especially as our knowledge of the microflora associated with health, disease and the transition from health to disease is minimal. It must also be recognised that the microflora associated with the transition state between health and disease may be different on different enamel surfaces (occlusal and approximal) and on exposed dentinal root surfaces.
Association and cause and effect
It is important to appreciate the difference between an association and cause and effect. For example, a study might show that a sample of the population which had a high incidence of skin cancer also consumed a greater than average quantity of ice cream. This does not mean that eating ice cream causes skin cancer; it merely shows an association, probably because people who expose their skin to sun radiation on the beach may also tend to eat more ice creams. A further example is that in the early part of the twentieth century, it was noticed that, when viewed over a measured period of time, the number of crimes increased with membership in the Church of England. This had nothing to do with criminals discovering religion. Rather, it was because both crimes and Church membership increased at the same time as the population increased. These two examples illustrate associations which are coincidental.
To establish />
Stay updated, free dental videos. Join our Telegram channel
VIDEdental - Online dental courses


