CHAPTER 3. Dental caries
Dental caries is progressive and, initially, subsurface demineralistion of teeth by bacterial acid. Caries is one of the most common of all diseases and still a major cause of loss of teeth.
The ultimate effect of caries is to break down enamel and dentine and thus open a path for bacteria to reach the pulp. The consequences are inflammation of the pulp and, later, of the periapical tissues. Acute pulpitis and apical periodontitis caused in this way are the most common causes of toothache. Infection can spread from the periapical region to the jaw and beyond. Though this is rare in Britain, people in other countries occasionally die from this cause.
AETIOLOGY
In 1890, W. D. Miller showed that lesions similar to dental caries could be produced by incubating teeth in saliva when carbohydrates were added. Miller concluded that caries could result from decalcification caused by bacterial acid production followed by invasion and destruction of any remaining tissue. Though he took a laudably cautious view as to how these findings should be interpreted, Miller’s basic hypothesis has been upheld, even though the infective nature of the disease was not confirmed until 1954 when Orland and his associates in the USA showed that caries did not develop in germ-free animals. However, dental caries develops only in the presence of several interacting variables (Fig. 3.1 and Box 3.1).
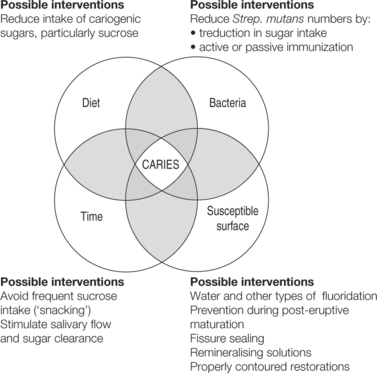 |
| Fig. 3.1
The four major factors in the aetiology of dental caries.
|
Box 3.1
Essential requirements for development of dental caries
1. Cariogenic (acidogenic) bacteria
2. Bacterial plaque
3. Stagnation areas
4. Fermentable bacterial substrate (sugar)
5. Susceptible tooth surfaces
6. Time
MICROBIOLOGY
Substantial evidence indicates that streptococci are essential for development of caries, particularly of smooth (interstitial) surfaces. These are viridans streptococci which are a heterogeneous group including Streptococcus mutans, S. sobrinus, S. salivarius, S. mitior and S. sanguis.
Viridans streptococci vary in their ability to attach to different types of tissues, their ability to ferment sugars (particularly sucrose) and the concentrations of acid thus produced. They also differ in the types of polysaccharides that they form.
Certain strains of S. mutans are strongly acidogenic; at low pH, with freely available sucrose, they also store an intracellular, glycogen-like, reserve polysaccharide. When the supply of substrate dries up, this reserve is metabolised to continue acid production for a time. Drastic reduction in dietary sucrose intake is followed by virtual elimination of S. mutans from plaque and reduces or abolishes caries activity. When sucrose is made freely available again S. mutans rapidly recolonises the plaque.
S. mutans strains are a major component of plaque in human mouths, particularly in persons with a high dietary sucrose intake and high caries activity (Fig. 3.2). S. mutans isolated from such mouths are virulently cariogenic when introduced into the mouths of animals.
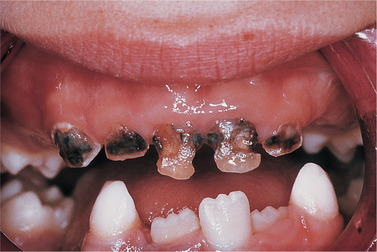 |
| Fig. 3.2
Extensive caries of decidous incisors and canines. This pattern of caries is particularly associated with the use of sweetened dummies and sweetened infant drinks.
|
However, simple clinical observation of the sites (interstitially and in pits and fissures) where dental caries is active, shows that the bacteria responsible are not those floating free in the saliva. Dental caries develops only at the interface between tooth surface and dental plaque in stagnation areas, particularly in occlusal fissures and approximally (Fig. 3.3).
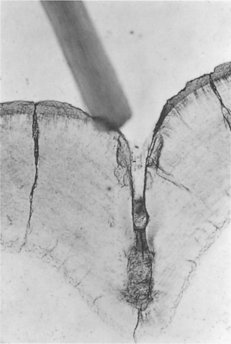 |
| Fig. 3.3
The stagnation area in an occlusal pit. A ground section of a molar showing the size of the stagnation area in comparison with a toothbrush bristle placed above it. The complete inaccessibility of the stagnation area to cleaning is obvious.
|
Bacterial polysaccharides
The ability of S. mutans to initiate smooth surface caries and form large amounts of adherent plaque depends on its ability to polymerise sucrose into high-molecular-weight, dextran-like, extracellular polysaccharides (glucans) (Box 3.2). The cariogenicity of S. mutans depends as much on its ability to form large amounts of insoluble extracellular glucans as on its ability to produce acid.
Box 3.2
Essential properties of cariogenic bacteria
• Acidogenic
• Able to produce a pH low enough (usually pH <5) to decalcify tooth substance
• Able to survive and continue to produce acid at low levels of pH
• Possess attachment mechanisms for firm adhesion to smooth tooth surfaces
• Able to produce adhesive, insoluble plaque polysaccharides (glucans)
Glucans enable streptococci to adhere to one another and to the tooth surface, probably via specific receptors. In this way, S. mutans and its glucans may initiate their attachment to the teeth and enable critical masses of plaque to be built up. Production of sticky, insoluble, extracellular glucan produced by strains of S. mutans is strongly related to their cariogenicity.
The importance of sucrose in this activity depends on the high energy of its glucose–fructose bond which allows the synthesis of polysaccharides by glucosyltransferase without any other source of energy. Sucrose is thus the main substrate for such polysaccharides. Other sugars are, to a variable degree, less cariogenic (in the absence of preformed plaque), partly because they are less readily formed into cariogenic glucans.
Plaque polysaccharides, synthesised by bacteria, play an essential role in the pathogenesis of dental caries. The proportions of the different types of polysaccharide, and the overall amounts formed, depend both on the kinds of bacteria present and the different sugars in the diet.
On a sucrose-rich diet, the main extracellular polysaccharides are glucans. Fructans formed from fructose are produced in smaller amounts. They are more soluble than glucans and less important in caries. Acid-producing microorganisms that do not produce insoluble polysaccharides do not appear to be able to cause caries of smooth surfaces. Even mutant strains of S. mutans which produce more soluble polysaccharides seem not to be cariogenic. Polysaccharides thus contribute to the adhesiveness, bulk and resistance to solution of plaque.
In the past, lactobacilli were thought to be the main cause of dental caries because they are numerous in the saliva and can be isolated from carious cavities. They are also acidogenic. However, they are present only in relatively small numbers in dental plaque until after caries has developed.
In gnotobiotic animals, lactobacilli are weakly cariogenic but some can produce fissure caries where adherent plaque formation is less important. Overall, there is little evidence that lactobacilli are clinically important in initiating dental caries, but they may contribute to tooth destruction after the process has started.
Many other microorganisms can also be found in plaque. The role of the many filamentous forms is not known. Strains of actinomyces are also found, particularly when caries is rampant, but appear capable only of causing root surface lesions.
Though bacteria are responsible for acid production, bacterial plaque (Fig. 3.4) enables large concentrations of them to adhere to the teeth and, in stagnation areas, prevents effective buffering of bacterial acids by saliva. Important points about microbiological aspects of dental caries are summarised in Box 3.3.
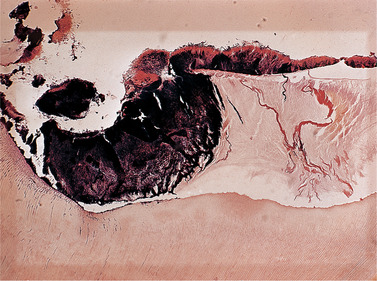 |
| Fig. 3.4
Bacterial plaque. A decalcified section showing darkly staining plaque lying on enamel and within a carious cavity. The plaque has remained intact and adherent to the enamel throughout the processes to which the specimen was subjected in preparation for sectioning.
|
Box 3.3
Microbiological aspects of dental caries
• Dental caries is a bacterial disease
• The organisms mainly responsible are specific strains of Streptococcus mutans
• The cariogenicity of S. mutans has been established by inoculating it into the mouths of otherwise germ-free animals (gnotobiotes)
• The presence of S. mutans in the human mouth is associated with caries activity
• Other bacteria including lactobacilli and other strains of streptococci are only weakly cariogenic or are non-cariogenic despite being able to produce acid
The cariogenicity of S. mutans depends on properties summarised in Box 3.4.
Box 3.4
Cariogenic properties of Streptococcus mutans
• It produces lactic acid from sucrose
• It can live at a pH as low as 4.2
• It forms large amounts of extracellular, sticky and insoluble glucan plaque matrix
• It adheres to pellicle and contributes to plaque formation
BACTERIAL PLAQUE
Plaque is a tenaciously adherent deposit that forms on tooth surfaces. It consists of an organic matrix containing a dense concentration of bacteria (Fig. 3.4, Fig. 3.5 and Fig. 3.6).
 |
| Fig. 3.5
This scanning electronmicrograph of plaque shows the large number of filamentous organisms and, in addition, many cocci clustered amongst them.
(By kind permission of Dr Sheila J Jones.)
|
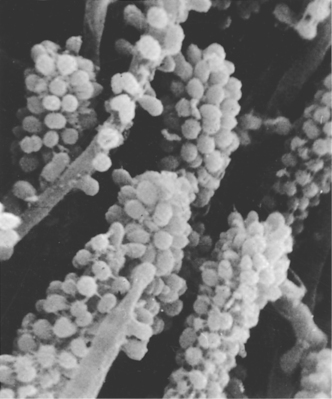 |
| Fig. 3.6
This scanning electronmicrograph at higher power shows cocci attached to filamentous organisms to produce the corn-cob type of arrangement sometimes seen in plaque.
(By kind permission of Dr Sheila J Jones.)
|
In microbiological terms, plaque is a biofilm. Biofilms consist of a hydrated viscous phase formed from bacteria and their extracellular polysaccharide matrices. In such a film, molecules and ions exist in concentrations that can be widely different from those of the surrounding fluid phase (saliva). Bacteria in biofilms can also exhibit cooperative activity and behave differently from the same species in isolation in a culture medium. As a consequence, a biofilm may be resistant to antimicrobials or to immunological defences to which the individual bacteria are normally sensitive. Bacterial plaque must therefore be regarded as a living entity and not as a mere collection of bacteria. In particular, the ability of dental plaque to concentrate and retain acid was recognised long before the special properties of biofilms were discovered.
Clinically, bacterial plaque is a tenaciously adherent deposit on the teeth. It resists the friction of food during mastication and can only be readily removed by toothbrushing. However, neither toothbrushing nor fibrous foods will remove plaque from inaccessible surfaces or pits (stagnation areas – see Fig. 3.3).
Plaque becomes visible, particularly on the labial surfaces of the incisors, when toothbrushing is stopped for 12–24 hours. It appears as a translucent film with a matt surface that dulls the otherwise smooth and shiny enamel. It can be made obvious when stained with disclosing agents. Little plaque forms under conditions of starvation, but it forms rapidly and abundantly on a high-sucrose diet.
In stagnation areas where they are undisturbed, plaque bacteria can form acid from sugars over sufficiently long periods as to attack tooth surfaces.
Adhesion of bacteria to the teeth from which they would otherwise be washed away is an essential requirement for the colonisation of enamel. Attachment depends on complex mechanisms and depends on such molecules as glucans and/or glucosyltransferase. Components of plaque, which act as adhesion receptors, include a group of proline-rich proteins (PRPs) from the saliva.
Stages of formation of bacterial plaque
If teeth are thoroughly cleaned by polishing with an abrasive, plaque quickly re-forms (Box 3.5).
Box 3.5
Stages of plaque formation
• Deposition of structureless cell-free, pellicle of salivary glycoprotein
• Further deposition of pellicle enhanced by bacterial action precipitating salivary proteins
• Colonisation of the cell-free layer by bacteria, particularly by S. sanguis and S. mutans strains within 24 hours
• Progressive build-up of plaque substance by bacterial polysaccharides
• Proliferation of filamentous and other bacteria as the plaque matures
Acid production in plaque
Sucrose diffuses rapidly into plaque and acid production quickly follows. These changes have been measured directly in the human mouth using microelectrodes in direct contact with plaque. It has been shown by this means that, after rinsing the mouth with a 10% glucose solution, the pH falls within 2–5 minutes, often to a level sufficient to decalcify enamel. Even though no more sucrose may be taken and the surplus is washed away by the saliva, the pH level remains at a low level for about 15–20 minutes; it returns only gradually to the resting level after about an hour. These changes (the so-called Stephan curve) are shown diagramatically in Figure 3.7. The rapidity with which the pH falls is a reflection of the speed with which sucrose can diffuse into plaque and the activity of the concentration of enzymes produced by the great numbers of bacteria in the plaque. The slow rate of recovery to the resting pH – a critical factor in caries production – depends mainly on factors summarised in Box 3.6.
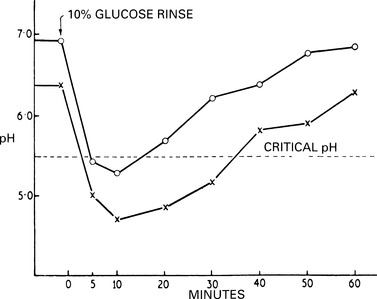 |
| Fig. 3.7
Stephan curves. This is the usual form of curves obtained when changes in reaction to bacterial plaque are measured directly. When the pH falls below the critical level, enamel may become demineralised. Patients with active caries tend to show a lower fall in pH, as in the lower curve. Noteworthy features are the very rapid fall in pH and the slow recovery to the normal level in spite of the very short time the sugar is in the mouth. Carbohydrates which are retained on the teeth have a more prolonged effect still.
|
Box 3.6
Factors contributing to maintenance of low plaque pH
• Rapid production of a high concentration of acid within the plaque temporarily overcomes local buffering
• Escape of acid into the saliva is delayed by the diffusion-limiting properties of plaque and its thickness
• Diffusion of salivary buffers into plaque hampered by the diffusion-limiting properties of plaque and its thickness
• Continued acid production from bacterial intracellular storage polysaccharides after dietary sugar is exhausted
Plaque minerals
In addition to bacteria and their polysaccharides, salivary components also contribute to the plaque matrix. Calcium, phosphorus and, often, fluorides are present in significant amounts. There is some evidence of an inverse relationship between calcium and phosphate levels in plaque and caries activity or sucrose intake. The ability of plaque to concentrate calcium and phosphorus is used in mineralising mouthwashes. The level of fluoride in plaque may be high, ranging from 15 to 75 ppm or more, and is largely dependent on the fluoride level in the drinking water and diet. This fluoride is probably mostly bound to organic material in the plaque but, at low pH levels, may become available and active in ionic form.
SUCROSE AS A PLAQUE SUBSTRATE
Direct measurement of pH changes in the mouth shows that there is intermittent acid production on the surface of the teeth and this follows the pattern shown earlier in the Stephan curves. Ingestion of sucrose leads to a burst of activity in the plaque so that the pH may fall low enough to attack enamel before slowly returning to the resting level. The frequency with which substrate is made available to the plaque is therefore important. When sucrose is taken as a sweet drink, any surplus beyond the capacity of the organisms in the plaque to metabolise at the time is washed away. If sucrose-containing drinks are taken repeatedly at short intervals, the supply of substrate to the bacteria can be sufficiently frequently renewed to cause acid in the plaque to remain persistently at a destructive level.
A similar effect may be caused by carbohydrate in sticky form, such as a caramel, which clings to the teeth and is slowly dissolved, releasing substrate over a long period. The effects of maintaining plaque activity by repeated administration of small quantities of sucrose have been demonstrated by the use of animal feeding devices to dispense metered quantities of sucrose in diets at fixed intervals. These show that a given amount of sucrose is more cariogenic when fed in small increments, but at intervals to maintain maximal plaque activity, than the same amount fed as a single dose.
Effects of sucrose on plaque polysaccharide production
As discussed earlier, the cariogenicity of plaque depends on its ability to adhere to the teeth, to resist dissolution by saliva and its protection of bacterial acids from salivary buffering. These properties depend on the formation of insoluble polysaccharides produced particularly by cariogenic strains of S. mutans.
Effects of sucrose on the oral microbial flora
Colonisation by cariogenic bacteria, especially S. mutans, is highly dependent on the sucrose content of the diet. In the absence of sucrose, S. mutans cannot usually be made to colonise the mouths of experimental animals. In humans, the plaque counts of S. mutans also appear to depend on the sucrose content of the diet. Severe reduction in dietary sucrose causes S. mutans to decline in numbers or disappear from plaque.
Experimental evidence for the essential contribution of sucrose to caries activity is summarised in Box 3.7.
Box 3.7
Experimental evidence for the critical contribution of sucrose to caries activity
• In caries-susceptible animals a sucrose-rich diet promotes caries production
• Caries is not induced in susceptible animals if sucrose is fed only by stomach-tube – its effect is entirely local
• Sucrose in sticky form clings to the teeth and remains available to bacteria for a longer period and is more cariogenic
• Sucrose-containing fluids are quickly cleared from the mouth and less cariogenic
• Frequent feeds of small quantities of sucrose are more cariogenic than the same total amount fed on a single occasion
• Desalivation, by delaying clearance of sugars, enhances caries activity
The importance of sucrose in human caries depends mainly on epidemiological studies and a few interventional studies. The findings of epidemiological studies are summarised in Box 3.8.
Box 3.8
Epidemiological evidence for sucrose as the cause of dental caries
• Low caries prevalence in populations with low sucrose intakes
• The decline in caries prevalence during wartime sucrose shortages
• The rise of caries prevalence with increasing availability of sucrose
• Archaeological evidence of low caries prevalence in eras before sucrose became freely available
• Low caries prevalence in disorders of sucrose metabolism (hereditary fructose intolerance)
In such studies, no association has been found between malnutrition and caries. Generally, the reverse is true and, when nutrition is poor, caries is infrequent. These diets also vary widely in content, from rice as the staple in China or coarsely ground cereals in Africa to a mainly meat and fish diet among Eskimos. The common feature of these diets, and one differentiating them sharply from westernised diets, is low or negligible consumption of refined sugar, particularly sucrose.
Britain is an example of a country where consumption of sucrose is exceptionally high and where the rest of the diet is more than adequate. A survey in England and Wales in 1973 showed, for example, that (in spite of the benefits of a National Health Service) 78% of 8-year-old children had active decay and many teeth which had been filled or lost from this cause. A survey in 1978 showed that dentate adults had on average only 13 sound and unfilled teeth. Though there has been dramatic improvement nationally, such consequences are still found in some populations.
In Britain and other countries, the incidence of caries has risen roughly parallel with rising consumption of sucrose. The incidence of caries has risen in spite of the much greater consumption of so-called ‘protective’ foods, namely dairy products, meat and fruit, in recent years. As a consequence of this more varied diet, carbohydrates overall have formed a smaller proportion of the diet. Nevertheless, sucrose forms a higher proportion of the carbohydrate component.
Where there has been a change from simple natural diets to a westernised diet, as in Alaska, caries has increased sharply and Inuit children now have a caries incidence at least as high as other American children. A similar state of affairs is developing in Africa as sweet eating and sweet between-meals ‘snacks’ have become popular. In Nigeria, many of the older adults are still totally caries-free, but the prevalence of dental caries among children and young people who have picked up sweet-eating habits is rising rapidly. This effect has also been strikingly well documented in the population of Tristan da Cunha who, up to the late 1930s, lived on a simple meat, fish and vegetable diet with a minimal sucrose content and had a very low caries prevalence. With the adoption of westernised diet, the caries prevalence had risen up to eightfold in some age groups by the mid-1960s. Even in Europe, in the 1930s and 1940s there were isolated communities, such as the Outer Hebrides or the Lötschental in Switzerland, where dental caries prevalence rose 20-fold or more when sucrose and sweets became widely consumed.
The effect of limiting sucrose consumption was shown on a vast scale during the Second World War. Those countries which suffered food shortages during the 1939–45 war had severe restrictions, mainly of meat, fats and sucrose. To maintain an adequate caloric intake, overall consumption of starchy carbohydrates rose considerably. In occupied Norway, where shortages were particularly severe, the pattern of dental caries during and after the war is shown diagrammatically in Figure 3.8. As sucrose became more plentiful at the end of the war, caries prevalence progressively rose.
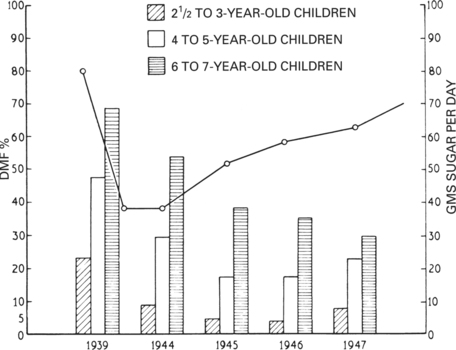 |
| Fig. 3.8
The wartime restrictions on diet and caries in Norway. The continuous line shows an estimate of the individual daily sugar consumption during and after the war. The heights of the columns show the incidence of caries in children of various ages. Rationing of sugar started in 1938, but it is apparent that the incidence of caries declined slowly and continued for a short time after sugar became freely available. The greatest reduction in caries was in the youngest group of children whose teeth were exposed to the wartime diet for the shortest periods; the mothers of these children, on the other hand, had been exposed to the wartime diet throughout pregnancy and for a long period before that.
(Mainly after Toverud G 1949 Journal of the American Dental Association 39:127.)
|
Caries has become epidemic only in relatively recent years as sucrose became cheaper and widely available. In Britain, there was a sudden, widespread rise in sucrose consumption in the middle of the 19th century. This resulted both from the falling cost of production and, in 1861, the abolition of a tax on sugar. Evidence from exhumed skulls confirms the low prevalence of caries before sucrose became widely available and the steady rise in prevalence thereafter.
Patients unable to metabolise fructose as a result of an enzyme deficiency cannot tolerate fructose-containing foods including disaccharides such as sucrose where fructose forms part of the molecule. These children therefore learn to avoid all sucrose-containing foods and have an unusually low incidence of caries.
Experimental studies on humans
In the Vipeholm study, over 400 adult patients were studied in a closed institution. They received a basic low-carbohydrate diet to establish a baseline of caries activity for each group. They were then divided into seven groups which were each allocated different diets. A control group received the basic diet made up to an adequate calorie intake with margarine. Two groups received supplements of sucrose at mealtimes, either in solution or as sweetened bread. The four remaining groups received sweets (toffees, caramels, or chocolate) which were eaten between meals.
The effects of sucrose in different quantities and of different degrees of adhesiveness, and of eating sucrose at different times were thus tested over a period of 5 years. Caries activity was greatly enhanced by the eating, between meals, of sticky sweets (toffees and caramels) that were retained on the teeth. Sucrose at mealtimes only had little effe/>
Stay updated, free dental videos. Join our Telegram channel
VIDEdental - Online dental courses


