Hormones and growth factors
Publisher Summary
This chapter discusses hormones and growth factors. Cells are highly sensitive to a variety of chemical signals, and the overall activities of tissues in different metabolic states are to a large extent controlled by hormones. Hormones are involved in both short- and long-term control. Hormones may be defined as substances that are produced in very small amounts and are carried in the blood from the site of their production to a specific target tissue or tissues where they bring about a response that is appropriate to the functioning of the body as a whole. Hormones are essential for normal development and activity, and most of them are produced continuously at a slow rate. Under certain conditions, however, much larger amounts are released and the factors that trigger their release are more or less specific for each endocrine gland. The effects of hormones are individual and striking. Hormones produced by specialized cells but not by clearly defined endocrine glands include gastrin, secretin, pancreozymin, and enterogastrone, which are produced in the alimentary canal, and renin and erythropoietin, which are produced by the kidney. Several other substances that have hormone-like activity are widely distributed and exert their effects in the area in which they are produced. Histamine, serotonin, and various plasma kinins are considered to be local hormones.
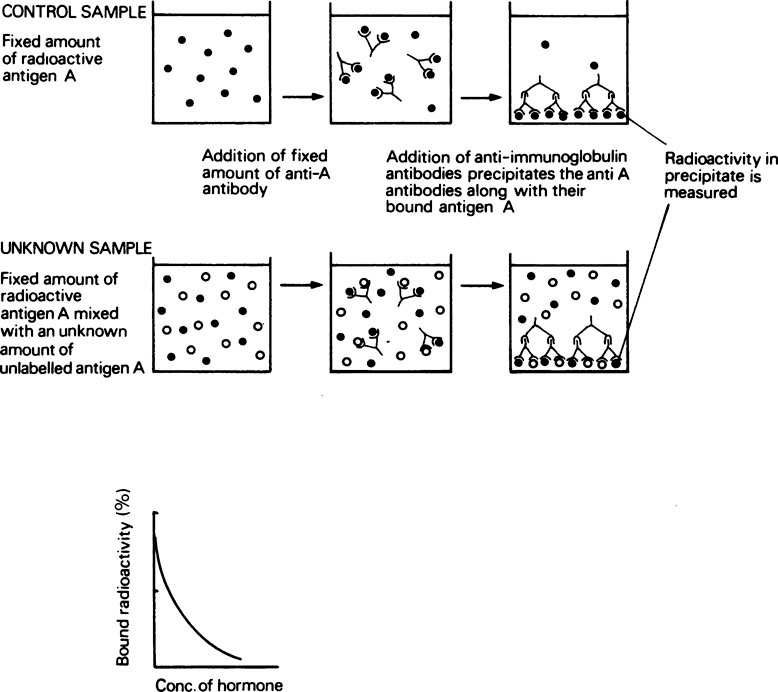
The determination of hormone concentrations by radioimmunoassay
In order to assay insulin by this method, a specific insulin-binding antibody must be produced. A sample of insulin is then radioactively labelled by incorporation of 125I into some of its tyrosine residues. When the radiolabelled hormone is mixed with the antibody, a complex is formed which can be separated by centrifugation, and the radioactivity that has been bound measured. The assay is calibrated by mixing a constant amount of radiolabelled insulin with known amounts of a standard solution of unlabelled insulin, adding an exact amount of antibody and measuring the radioactivity bound. Since the antibody will bind native insulin and iodinated insulin with the same affinity, there will be competition between labelled and unlabelled insulin for the available binding sites. The more unlabelled insulin there is present, the less labelled insulin will be bound and vice versa. A typical calibration curve for such an assay is shown in Figure 24.1. The concentration of insulin in an unknown solution may then be measured by mixing the solution with radioactive insulin and antibody as before, measuring the radioactivity bound and reading the insulin concentration from the calibration curve.
The mode of action of hormones
The actions of hormones are very diverse. Some, e.g. glucagon, have specific and clearly defined effects on the metabolism of particular tissues. Others such as growth hormone and the sex hormones have more general effects on growth and development while the tropic hormones stimulate their target tissues to secrete further hormones. For example the adrenocorticotropic hormone (ACTH), which is produced in the anterior pituitary, promotes the synthesis and release of glucocorticoids by the adrenal cortex. A list of some of the hormones and their effects on metabolism is given in Table 24.1.
Table 24.1
Hormones and their effects on metabolism
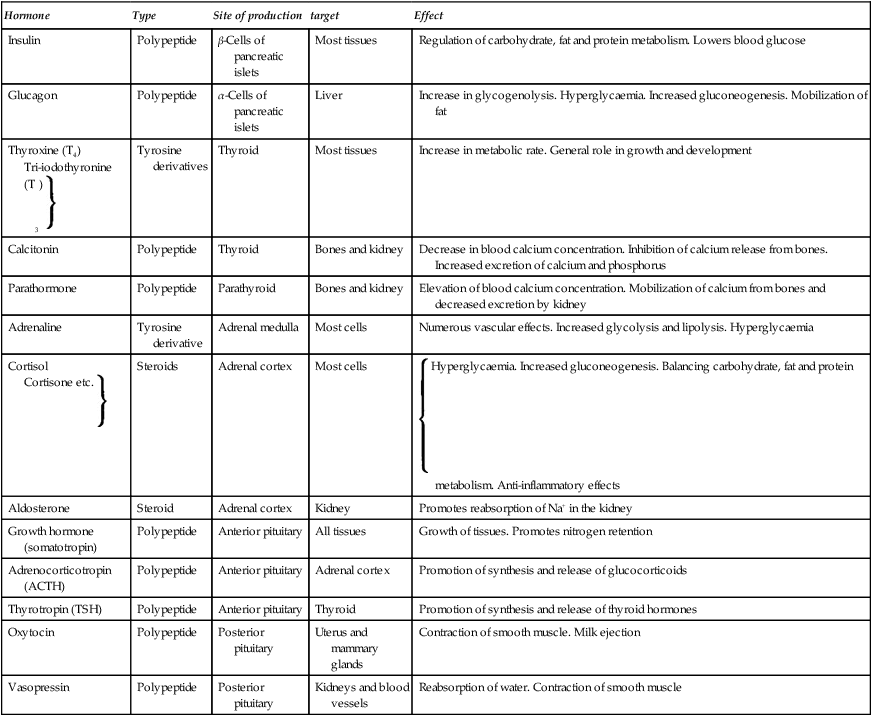
| Hormone | Type | Site of production | target | Effect |
| Insulin | Polypeptide | β-Cells of pancreatic islets | Most tissues | Regulation of carbohydrate, fat and protein metabolism. Lowers blood glucose |
| Glucagon | Polypeptide | α-Cells of pancreatic islets | Liver | Increase in glycogenolysis. Hyperglycaemia. Increased gluconeogenesis. Mobilization of fat |
| Thyroxine (T4) Tri-iodothyronine (T3) 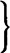 |
Tyrosine derivatives | Thyroid | Most tissues | Increase in metabolic rate. General role in growth and development |
| Calcitonin | Polypeptide | Thyroid | Bones and kidney | Decrease in blood calcium concentration. Inhibition of calcium release from bones. Increased excretion of calcium and phosphorus |
| Parathormone | Polypeptide | Parathyroid | Bones and kidney | Elevation of blood calcium concentration. Mobilization of calcium from bones and decreased excretion by kidney |
| Adrenaline | Tyrosine derivative | Adrenal medulla | Most cells | Numerous vascular effects. Increased glycolysis and lipolysis. Hyperglycaemia |
| Cortisol Cortisone etc. 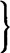 |
Steroids | Adrenal cortex | Most cells | 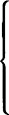 Hyperglycaemia. Increased gluconeogenesis. Balancing carbohydrate, fat and protein metabolism. Anti-inflammatory effects Hyperglycaemia. Increased gluconeogenesis. Balancing carbohydrate, fat and protein metabolism. Anti-inflammatory effects |
| Aldosterone | Steroid | Adrenal cortex | Kidney | Promotes reabsorption of Na+ in the kidney |
| Growth hormone (somatotropin) | Polypeptide | Anterior pituitary | All tissues | Growth of tissues. Promotes nitrogen retention |
| Adrenocorticotropin (ACTH) | Polypeptide | Anterior pituitary | Adrenal cortex | Promotion of synthesis and release of glucocorticoids |
| Thyrotropin (TSH) | Polypeptide | Anterior pituitary | Thyroid | Promotion of synthesis and release of thyroid hormones |
| Oxytocin | Polypeptide | Posterior pituitary | Uterus and mammary glands | Contraction of smooth muscle. Milk ejection |
| Vasopressin | Polypeptide | Posterior pituitary | Kidneys and blood vessels | Reabsorption of water. Contraction of smooth muscle |
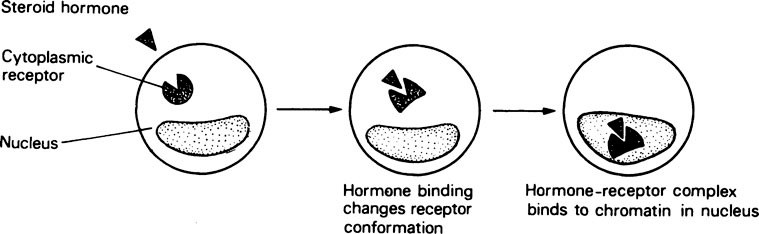
Steroid hormones
After entering the target cell, each type of steroid hormone binds with high affinity to a specific type of receptor in the cytoplasm. The hormone-receptor complex then undergoes some sort of transformation, as shown by an increase in its sedimentation rate, after which it migrates to the nucleus and binds to the DNA at specific sites (Figure 24.2). After binding to the chromatin the hormone-receptor complex seems to stimulate the transcripton of specific genes with the production of the corresponding mRNA molecules. The response to a steroid hormone may take place in two stages. In the primary response there is a direct induction of transcription of certain genes while, in the secondary response, further genes are activated by the products of the primary response. The actual nature of the response is determined by the nature of the target cell. For example, oestradiol causes epithelial development in the uterus and mammary glands while it decreases the resorption of bone.
The water-soluble hormones
The majority of hormones that bind to cell surface receptors do not enter the cell at all but instead they generate an intracellular signal or second messenger and it is this which is responsible for altering the behaviour of the cell. They may do this by changing the activity of an enzyme bound to the plasma membrane or by opening or closing gated ion channels (page 197) within the plasma membrane. The most important membrane-bound enzyme with respect to such hormones is adenylate cyclase which catalyses the synthesis of cAMP from ATP in the cytoplasm.
Stay updated, free dental videos. Join our Telegram channel
VIDEdental - Online dental courses


